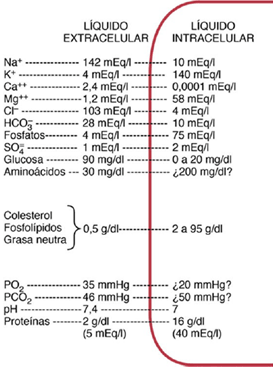
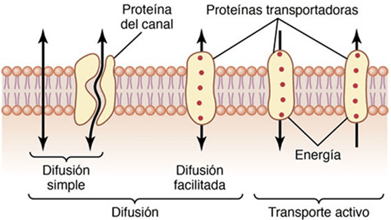
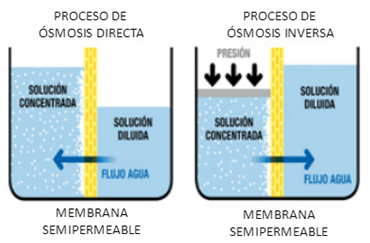
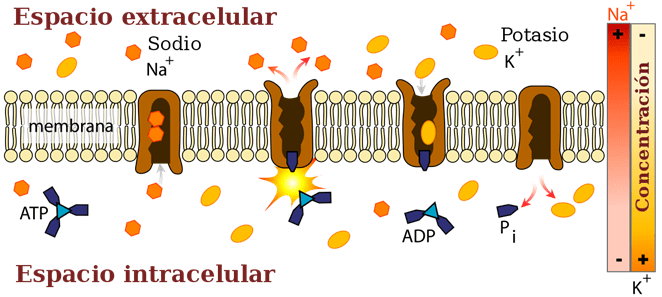
CLASIFICACIÓN DE LA SENSIBILIDAD SOMÁTICA
l. La sensibilidad mecanorreceptora abarca las sensibilidades táctil y posicional (propioceptiva).
2. La sensibilidad termorreceptora detecta los aumentos o descensos de temperatura.
3. La sensibilidad al dolor detecta el daño tisular o la liberación de moléculas específicas mediadoras del dolor.
Las modalidades sensitivas vehiculadas por los sistemas sensitivos somáticos son el tacto discriminatorio (localizado de manera poco precisa), la presión, la vibración y las sensaciones de posición estática y velocidad de movimiento, que se denominan en conjunto propiocepcián. Las sensaciones exteroceptivas son aquellas que se originan por la estimulación de las estructuras de la superficie corporal, como la piel y el tejido subcutáneo, o más profundas, como los músculos, las fascias y los tendones. En cambio, las señales sensitivas que nacen de los órganos internos (estructuras derivadas del endodermo) se denominan viscerales.
Detección y transmisión de las sensaciones táctiles
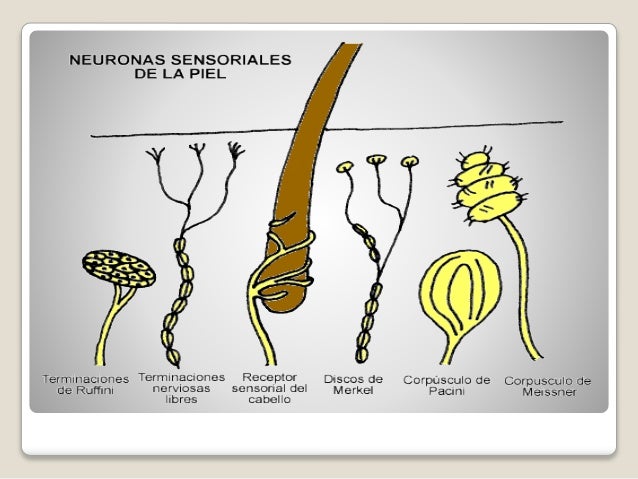
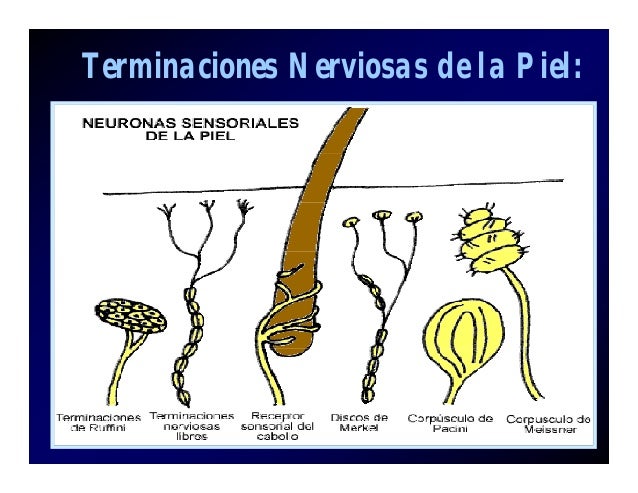
Aunque el tacto, la presión y la vibración suelen clasificarse como sensibilidades separadas y diferentes, su detección corresponde a la misma categoría general de receptores táctiles: los mecanorreceptores. Son seis, al menos, los tipos de mecanorreceptores clasificados como receptores táctiles:
- Las terminaciones nerviosas libres se encuentran, en un número variable, en todas las regiones de la piel y en la córnea del ojo.
- El corpúsculo de Meissner es un receptor de adaptación rápida, encapsulado, situado en las regiones lampiñas (glabras) de la piel, como las yemas de los dedos y los labios, zonas especialmente sensibles a la estimulación táctil más ligera.
- Los discos de Merkel (conocidos como receptores de terminación bulbar) se encuentran en la piel lampiña, pero también, en cantidad moderada, en la superficie con pelo.Estos receptores son de adaptación relativamente lenta y al parecer median en el contacto continuo de los objetos contra la piel.
- Los órganos terminales del pelo (terminacionesperítricas) están entremezclados en la base de cada pelo de la superficie corporal, son receptores de adaptación rápida y detectan el movimiento de los objetos sobre la superficie de la piel que desplazan los pelos.
- Las terminaciones de Ruffini son terminaciones encapsula das de la piel y tejidos más profundos, así como de las cápsulas articulares, con escasa adaptación, que detectan el tacto y la presión continuos aplicados a la piel o los movimientos alrededor de la articulación donde se ubican.
- Los corpúsculos de Pacini están presentes en la piel y en tejidos más profundos, como las fascias, se adaptan con rapidez y, en principio, resultan muy importantes para detectar la vibración y otros cambios rápidos en el estado mecánico de los tejidos.
La mayoría de estas categorías de receptores táctiles trans miten señales por fibras mielínicas relativamente grandes, con una velocidad de conducción rápida. En cambio, las terminaciones nerviosas libres se asocian con pequeñas fibras mielínicas y fibras amielínicas de tipo C, que conducen con una velocidad relativamente baja.
Cada uno de los receptores táctiles participa también en la detección de vibraciones. Los corpúsculos de Paccini detectan los estímulos vibratorios más rápidos (de 30 a 800 ciclos por segundo) y se asocian con fibras mielínicas grandes, de conducción rápida. La vibración de baja frecuencia (hasta 80 ciclos por segundo) estimula los corpúsculos de Meissner y otros receptores táctiles que suelen transmitir con velocida des bastante bajas y cuya adaptación es menos rápida que la de los corpúsculos de Paccini.
La sensación de cosquilleo o picor se relaciona con terminaciones nerviosas libres muy sensibles y de adaptación rápida, situadas en las capas superficiales de la piel, que se transmiten sobre todo por fibras de tipo C. La función de esta modalidad sensitiva es, con toda probabilidad, llamar la atención a las irritaciones cutáneas leves que se alivian con el movimiento o el raspado, estímulo que, en principio, anula las señales de picor.
Vías sensitivas para la transmisión de señales somáticas en el sistema nervioso central
Las vías principales para la transmisión de las señales somatosensitivas son el sistema de la columna dorsal-lemnisco medial y el sistema anterolateral. Con escasas excepciones, la información sensitiva transportada por las fibras nerviosas de la superficie corporal (con excepción de la cara) entra en la médula espinal por las raíces dorsales. Una vez dentro del sistema nervioso central, la señal se segrega por una de dos vías. Las señales originadas en los termorreceptores y receptores para el dolor son procesadas por el sistema anterolateral y las que nacen en los meca norreceptores discurren por el sistema de la columna dorsal lemnisco medial (CD-LM), que vehicula modalidades como el tacto discriminatorio, la vibración y la propiocepción. De manera análoga, la información somatosensitiva de la cara es canalizada fundamentalmente por ramos del nervio trigémino; cuando estas fibras entran en el tronco encefálico, también se segregan en dos vías: una especializada en el procesamiento del dolor, la temperatura y el tacto grosero, y la otra responsable del tacto discriminatorio, las vibraciones y la propiocepción.
Transmisión por el sistema de la columna dorsal-lemnisco medial
La anatomía del sistema de la columna dorsal-lemnisco medial se caracteriza por un alto grado de organización somatotópica (espacial)
· Neuronas sensitivas primarias. Las prolongaciones centrales de las neuronas sensitivas primarias del ganglio raquídeo que entran en la médula espinal por la división medial de la zona de entrada en la raíz dorsal son las fibras mie!ínicas mayores, que portan las señales relacionadas con el tacto discriminatorio, la vibración y la propiocepción. A su entrada en la médula, algunas de estas fibras entablan sinapsis locales en la sustancia gris, mientras que muchas se dirigen sencillamente hasta la zona de la columna dorsal y ascienden, sin ningún tipo de sinapsis, hasta alcanzar los núcleos de la columna dorsal de la parte inferior del bulbo. Aquí, las fibras que vehiculan la información de los miembros inferiores establecen sinapsis con el núcleo grácil, mientras que las del miembro superior terminan en el núcleo cuneiforme.
• Núcleos de la columna dorsal. Los axones de las células de los núcleos cuneiforme y grácil forman el lemnisco medial, que atraviesa la línea media por la parte inferior del bulbo, a modo de decusación sensitiva. Este haz de fibras continúa rostralmente hasta el tálamo, donde los axones terminan en el complejo ventrobasal, principalmente en el núcleo ven trolateral posterior (VPL). Los axones de las neuronas VPL penetran después en la rama posterior de la cápsula interna y se proyectan hacia el área somatosensitiva I en la circunvolución poscentral.
• Vía del lemnisco medial.
Las fibras del sistema de la columna dorsal-lemnisco medial poseen un alto grado de organización somatotópica (orientación espacial). Las fibras que vehiculan las señales del miembro inferior ascienden por la porción medial de la columna dorsal, terminan en el núcleo grácil y forman la porción ventral y lateral del lemnisco medial. Estas fibras acaban termi nando lateralmente en las neuronas VPL, que proyectan hacia la porción más medial del área somatosensitiva I, en la pared medial del hemisferio. La información del miembro superior discurre por la parte lateral de la columna dorsal, termina en el núcleo cuneiforme y entra en la porción dorsal y medial del lemnisco medial. Estas fibras establecen sinapsis en la porción medial de las neuronas VPL y alcanzan finalmente el territorio del área somatosensitiva I para el miembro superior en el hemisferio contralateral a la superficie del cuerpo originaria de las señales.
A lo largo de todo este sistema existe una relación, punto a punto, entre el origen periférico y la terminación en el área somatosensitiva I.
• Señales somatosensitivas de la cara.
Las señales somatosensitivas táctiles de la cara viajan por el nervio trigémino y entran en el tronco encefálico a la altura central de la protuberancia, donde las fibras sensitivas primarias terminan en el núcleo sensitivo principal del trigémino. Desde aquí, los axones atraviesan la línea media y siguen un curso rostral, adyacente al lemnisco medial, para terminar finalmente en posición medial en una parte del complejo ventrobasal, el núcleo ventral posteromedial (VPM). Este sistema de fibrases comparable al de la columna dorsal-lemnisco medial y transporta una información somatosensitiva similar de la cara.
Áreas somatosensitivas de la corteza cerebral.
La circunvolución poscentral comprende la corteza somatosensitiva primaria, que corresponde a las áreas de Brodmann 3, 1 y 2. El área somatosensitiva II es mucho más pequeña que el área I y se encuentra justo posterior a la región facial del área I, bordeando la cisura lateral. Dentro del área somatosensitiva I, se mantiene la segregación de las regiones corporales, de modo que la cara ocupa una posición ventral, la más próxima a la cisura de Silvia, el miembro superior continúa medial y dorsalmente desde la región facial y se extiende hacia la convexidad del hemisferio, y el miembro inferior se proyecta sobre la cara medial del hemisferio. De hecho, las áreas 3, 1 y 2 contienen una representación completa, pero separada, del cuerpo. Dentro de cada una de estas representaciones corporales hay un volumen desigual de la corteza dedicado a cada región. Las superficies corporales con una elevada densidad de receptores sensitivos están representadas por áreas mayores en la corteza que aquellas con una densidad relativamente baja de receptores.
Anatomía funcional de la corteza somatosensitiva primaria
• Contiene seis capas celulares, con disposición horizontal, numeradas de I a VI, empezando por la capa I de la superficie cortical. La más característica es la capa IV porque recibe importantes proyecciones de los núcleos VPL y VPM del tálamo ventrobasal. Desde aquí, la información se propaga dorsalmente hasta las capas I a llI, y ventralmente hasta las capas V y VI.
• Contiene un ejército de co lumnas neuronales, dispuestas verticalmente, que se extiende por las seis capas. Se trata de columnas con una determinación funcional, cuya anchura varía entre 0,3 y 0,5 mm; cada una contiene aproximadamente 10.000 neuronas, según los cálculos. En la porción más anterior del área somatosensitiva I 3, la disposición vertical de las columnas se ocupa de las aferencias musculares, mientras que la parte posterior de esa área procesa las señales cutáneas de entrada. Las columnas verticales del área 1 procesan información cutánea adicional y las del área 2 se ocupan de la presión y de la propiocepcion.
Las funciones de las áreas somatosensitiva primaria y de asociación se pueden inferir a partir del estudio de pacientes con lesiones en estas zonas
• Las lesiones que dañan la corteza somatosensitiva primaria producen: 1) incapacidad para localizar de manera precisa los estímulos cutáneos en la superficie corporal, a pesar de que se conserva cierta capacidad de localización aproximada; 2) incapacidad para valorar el grado de presión o el peso de los objetos que contactan con la piel, y 3) incapaci dad para reconocer los objetos por el tacto o la textura (astereognosia).
• Las lesiones que dañan las áreas de Brodmann 5 ,7 y afectan a la corteza de asociación para la sensibilidad somática.
Los signos y síntomas habituales abarcan: 1) incapacidad para reconocer objetos con una forma o textura relativa mente compleja cuando se palpan con la mano contralateral; 2) pérdida de la conciencia del lado contralateral (heminegligencia) (este síntoma resulta más agudo con las lesiones del lóbulo parietal no dominante), y 3) al palpar un objeto, el paciente solo reconoce el lado ipsolateral a su lesión y hace caso omiso del contralateral.

Características generales de la transmisión y el análisis de las señales en el sistema de la columna dorsallemnisco medial
El campo receptor de una neurona cortical del área sormatosensitiva I depende de la de neuronas sensitivas primarias, neuronas de los núcleos de la columna dorsal y neuronas talámicas que emiten proyecciones aferentes hacia la neurona del área I.
La distinción entre dos puntos permite explorar el sistema de la columna dorsal-lemnisco medial. Este método suele utilizarse para averiguar la capacidad de una persona para discriminar entre dos estímulos cutáneos, aplicados simultáneamente, como dos «puntos» diferentes (distinción entre dos puntos). Esta capacidad varía mucho a lo largo de la superficie corporal. Si se aplican simultáneamente dos estímulos, con una proximidad de 1 a 2 mm, sobre las yemas de los dedos y los labios, se pueden diferenciar como tales, pero en la espalda se necesita una separación mínima de 30 a 70 mm para su discriminación. Esta función depende de elementos de procesamiento central situados en la vía de la columna dorsal-lemnisco medial que reconocen, como distintas y no superpuestas, dos señales excitadoras generadas en la periferia.
La inhibición lateral es el mecanismo que utiliza el sistema nervioso para «afinar» la transmisión de las señales.
Este proceso se vale de la inhibición de las aferencias de una porción periférica de un campo receptivo para definir mejor los límites de la zona excitada. En el sistema de la columna dorsal-lemnisco medial, la inhibición lateral ocurre en los núcleos de la columna dorsal y en los del tálamo.
El sistema de la columna dorsal-lemnisco medial detecta con especial eficacia los estímulos rápidamente cambiantes y repetitivos, que constituyen la base de la sensibilidad vibratoria. Esta capacidad reside en los corpúsculos de Paccini de adaptación rápida, que detectan vibraciones de hasta 700 ciclos por segundo, y en los corpúsculos de Meissner, que detectan frecuencias algo más bajas, del orden de 200 ciclos por segundo o menos.
La conciencia de combinación o del movimiento corporal se conoce como sensibilidad propiorreceptora. La sensación de movimiento corporal se conoce también como cinestesia o propiorrecepción dinámica. El sistema nervioso se sirve deuna combinación de receptores táctiles, musculares y de las posición propiorreceptora. Se cree que los receptores táctiles de la piel y de las cápsulas articulares resultan determinantes en la emisión de señales propiorreceptoras de movimientos de pequeñas regiones corporales, como los dedos de la mano.
Cuando se realizan movimientos complejos de los miembros superiores o inferiores, con aumento de algunos ángulos articulares y disminución de otros, los husos musculares contribuyen decisivamente a la sensibilidad posicional. Si la angulación de la articulación es extrema, el estiramiento impuesto a los ligamentos y tejidos profundos que rodean la articulación activa los corpúsculos de Paccini y las terminaciones de Ruffini. Estas últimas, receptores de adaptación rápida, probablemente detecten la velocidad de cambio del movimiento.
Transmisión de señales sensitivas menos esenciales por la vía anterolateral
Las señales que viajan por las pequeñas fibras mielínicas y las fibras amielínicas C pueden provenir de los receptores táctiles (de ordinario, terminaciones nerviosas libres) de la piel. Esta información se transmite, junto con las señales de dolor y de temperatura, por la porción anterolateral de la sustancia blanca de la médula espinal. El sistema anterolateral se extiende hasta la parte ventrobasal del tálamo y los núcleos talámicos intralaminar y posterior. Pese a que algunos estímulos dolorosos se localizan con bastante precisión, la organización precisa, punto a punto, del sistema de la columna dorsal-lemnisco medial y la organización relativamente difusa del sistema anterolateral explican probablemente la capacidad localizadora menos efectiva de este último.
Las características de transmisión por la vía anterolateral se parecen a las de la columna dorsal-lemnisco medial, con estas diferencias: 1) la velocidad de transmisión se reduce entre la mitad y un tercio de la de la columna dorsal-lemnisco medial; 2) el grado de localización espacial es escaso; 3) la graduación de la intensidad es mucho menos acusada, y 4) la capacidad de transmitir señales rápidas y repetidas es escasa. Aparte del dolor y de la temperatura, este sistema transmite las sensaciones de cosquilleo y picor, el tacto grosero y las sensaciones sexuales.