Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco. Algunos de los principios básicos de la contracción se aplican también a los diferentes tipos de músculos

Anatomía fisiológica del musculo esquelético
En la mayoria de los músculos esqueléticos las fibras se extienden a lo largo de toda la longitud del musculo. Cada fibra esta inervada por una sola terminación nerviosa.
Las miofibrillas están formadas por filamentos de actina y miosina. Cada fibra muscular contiene varios cientos a varios miles de miofibrillas. Cada miofibrilla está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular. Los filamentos gruesos son miosina y los delgados actina:
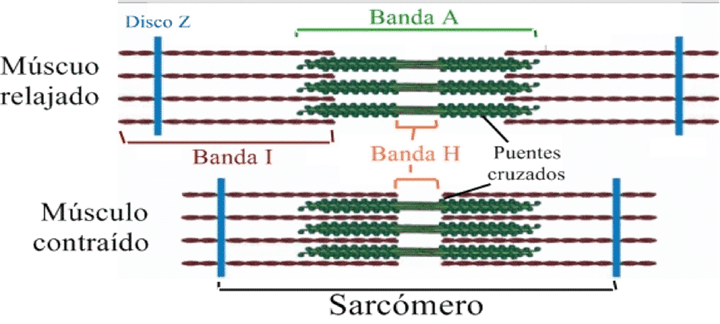
- Bandas claras y oscuras: las bandas claras contienen solo filamentos de actina y se denominan bandas I. Las bandas oscuras, llamadas bandas A, contienen filamentos de miosina asi como los extremos de los filamentos de actina. La longitud de la banda A es la longitud del filamento del filamento de miosina. La longitud de la banda I cambia con la contracción muscular.
- Puentes cruzados: Pequeñas proyecciones que se originan en los lados de los filamentos de miosina, la interacción entre estos y los filamentos de actina produce la contracción.
- Disco Z: los extremos de los filamentos de actina están unidos a los discos Z. El disco Z pasa a través de las miofibrillas y de una a otras, uniéndolas y alineándolas a lo largo de la fibra muscular. Por tanto, toda la fibra muscular tiene bandas claras y oscuras, dando al musculo esquelético y cardiaco un aspecto estriado.
- Sarcómero: porción de la miofibrilla que está entre dos discos Z sucesivos.
Mecanismo general de la contracción muscular
El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales:
1. Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
3. La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
6. El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.
7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
Mecanismo molecular de la contracción muscular
La contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. Las fuerzas mecánicas que se generan por las interacciones entre la actina y los filamentos de miosina hacen que los filamentos de actina últimos deslicen hacia dentro entre los filamentos de miosina. En el estado relajado, estas fuerzas están inactivas, pero cuando un potencial de acción viaja a lo largo de la fibra muscular hace que el retículo sarcoplasmático libere grandes cantidades de iones calcio que activan las fuerzas de atracción entre los filamentos de miosina y de actina, y origina el inicio de la contracción.
La molécula de miosina está formada por seis cadenas polipeptídicas, dos cadenas pesadas, y cuatro cadenas ligeras. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular. Los brazos y las cabezas que protruyen se denominan en conjunto puentes cruzados. Cada puente cruzado es flexible en dos puntos denominados bisagras, una en el punto en el que el brazo sale del cuerpo del filamento de miosina y la otra en el punto en el que la cabeza se une al brazo. Los brazos articulados permiten que las cabezas se separen del cuerpo del filamento de miosina o que se aproximen a este. Las cabezas articuladas, a su vez, participan en el proceso real de contracción. El filamento de miosina está enrollado de modo que cada par sucesivo de puentes cruzados está desplazado en sentido axial 120° con respecto al par previo. Esta torsión garantiza que los puentes cruzados se extiendan en todas las direcciones alrededor del filamento.
Interacción de un filamento de miosina, dos filamentos de actina y los iones calcio para producir la contracción
- Inhibición del filamento de actina por el complejo troponina-tropomiosina: un filamento de actina puro sin la presencia del complejo troponina-tropomiosina se une instantánea e intensamente a las cabezas de las moléculas de miosina. Después, si se añade el complejo troponina-tropomiosina al filamento de actina, no se produce la unión entre la miosina y la actina. En consecuencia, estos puntos no se pueden unir a las cabezas de los filamentos de miosina para producir la contracción. Antes de que se produzca la contracción, se debe inhibir el efecto bloqueante del complejo troponina-tropomiosina.
- Activación del filamento de actina por iones calcio: en presencia de grandes cantidades de iones calcio, se inhibe el propio efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina. No se conoce el mecanismo de esta inhibición, aunque una hipótesis es la siguiente: cuando los iones calcio se combinan con la troponina C, de la que una molécula se puede unir intensamente con hasta cuatro iones calcio, el complejo de troponina probablemente experimenta un cambio conformacional que en cierto modo tira de la molécula de tropomiosina y la desplaza hacia zonas más profundas del surco que hay entre las dos hebras de actina. Esta acción «descubre» los puntos activos de la actina, permitiendo de esta manera que atraigan a las cabezas del puente cruzado de miosina y que produzcan la contracción
- Interacción entre el filamento de actina «activado» y los puentes cruzados de miosina: teoría de la «cremallera» de la contracción: tan pronto como el filamento de actina es activado por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y de algún modo esto hace que se produzca la contracción. Aunque el mecanismo preciso mediante el que esta interacción entre los puentes cruzados y la actina produce la contracción sigue siendo en parte teórico, una hipótesis para la que hay datos considerables es la teoría de la «cremallera» (o teoría del «trinquete») de la contracción.
Características de la contracción de todo el musculo
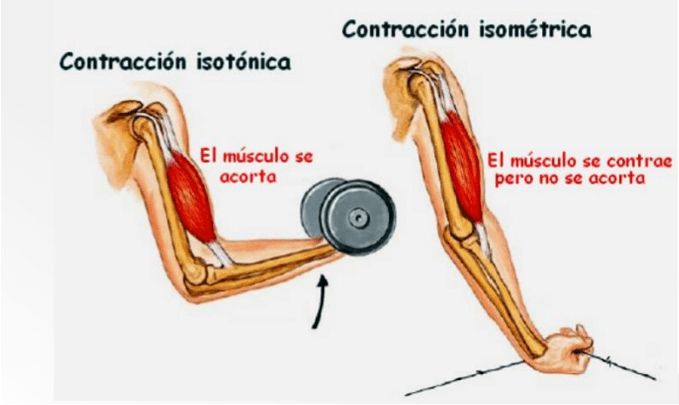
Las contracciones isométricas no acortan el musculo, mientras que las contracciones isotónicas lo acortan a una tensión constante.
Se dice que la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción.
Fibras musculares rápidas frente a lentas:
Fibras lentas (tipo I, músculo rojo):
- Las fibras son más pequeñas que las fibras rápidas.
- Las fibras lentas están también inervadas por fibras nerviosas más pequeñas.
- En comparación con las fibras rápidas, las fibras lentas tienen un sistema de vascularización más extenso y más capilares para aportar cantidades adicionales de oxígeno.
- Las fibras lentas tienen números muy elevados de mitocondrias, también para mantener niveles elevados de metabolismo oxidativo.
- Las fibras lentas contienen grandes cantidades de mioglobina, una proteína que contiene hierro y que es similar a la hemoglobina de los eritrocitos. La mioglobina se combina con el oxígeno y lo almacena hasta que sea necesario, lo cual acelera también notablemente el transporte de oxígeno hacia las mitocondrias. La mioglobina da al músculo lento un aspecto rojizo y el nombre de músculo rojo.
Fibras rápidas (tipo II, músculo blanco):
- Las fibras rápidas son grandes para obtener una gran fuerza de contracción.
- Existe un retículo sarcoplásmico extenso para una liberación rápida de iones calcio al objeto de iniciar la contracción.
- Están presentes grandes cantidades de enzimas glucolíticas para la liberación rápida de energía por el proceso glucolítico.
- Las fibras rápidas tienen una vascularización menos extensa que las lentas, porque el metabolismo oxidativo tiene una importancia secundaria.
- Las fibras rápidas tienen menos mitocondrias que las lentas, también porque el metabolismo oxidativo es secundario. Un déficit de mioglobina roja en el músculo rápido le da el nombre de músculo blanco.
Mecánica de contracción del musculo esquelético
Sumación significa la adición de los espasmos individuales para aumentar la intensidad de la contracción muscular global. La sumación se produce de dos maneras:
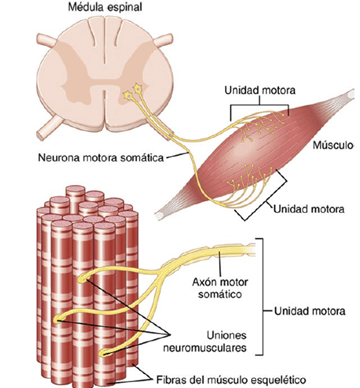
- Sumación de fibras múltiples: cuando el sistema nervioso central envía una señal débil para contraer un músculo, las unidades motoras más pequeñas del músculo se pueden estimular con preferencia a las unidades motoras de mayor tamaño. Después, a medida que aumenta la intensidad de la señal, también se empiezan a excitar unidades motoras cada vez mayores, de modo que las unidades motoras de mayor tamaño con frecuencia tienen una fuerza contráctil hasta 50 veces mayor que las unidades más pequeñas. Este fenómeno, denominado principio de tamaño, es importante porque permite que se produzcan gradaciones de la fuerza muscular durante la contracción débil en escalones pequeños, mientras que los escalones se hacen cada vez mayores cuando son necesarias grandes cantidades de fuerza.
- Sumación de frecuencia y tetanización: a medida que aumenta la frecuencia, se llega a un punto en el que cada nueva contracción se produce antes de que haya finalizado la anterior. En consecuencia, la segunda contracción se suma parcialmente a la primera, de modo que la fuerza total de la contracción aumenta progresivamente al aumentar la frecuencia. Cuando la frecuencia alcanza un nivel crítico, las contracciones sucesivas finalmente se hacen tan rápidas que se fusionan entre sí, y la contracción del músculo entero parece ser completamente suave y continua, como se muestra en la figura. Este proceso se denomina tetanización.
