La corteza cerebral se compone de una fina capa de neuronas, con un grosor de 2 a 5 mm y una superficie total cercana a 0.25 m²; en la corteza hay 100,000 millones de neuronas.
La mayoría de las neuronas corticales pertenecen a una de estas tres categorías:
- Granulares (o estrellladas).
- Fusiformes.
- Piramidales.
Las células granulares son neuronas de axones cortos, con circuitos locales, que utilizan glutamato (excitador) GABA (inhibidor) como neurotransmisores.
Las células fusiformes proyectan al tálamo, mientras que las neuronas piramidales lo hacen a otros lugares del hemisferio ipsolateral o contralateral y a una serie de estructuras subcorticales, como el núcleo rojo, la base de la protuberancia y la médula espinal.
Las neuronas de la corteza cerebral se organizan en seis capas horizontales. La capa IV recibe señales sensitivas aferentes del tálamo, mientras que las neuronas de la capa V dan proyecciones subcorticales largas para el tronco del encéfalo y la médula espinal.
Las fibras córticotalámica se originan en las en las células de la capa VI.
Las capas I, II Y III están especializadas en recibir aferencias y emitir eferencias a otras partes de la corteza del mismo hemisferio o del contralateral.

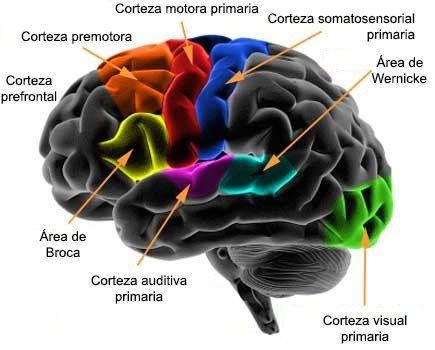
Se ha probado claramente mediante estudios que muchas áreas de la corteza cerebral se especializan en funciones concretas. Algunas áreas, denominadas corteza primaria, presentan conexiones directas con la médula espinal para controlar el movimiento, y otras regiones primarias reciben aferencias sensitivas de diversos núcleos del tálamo que representan cada uno de los sentidos especiales (excepto el olfato) y la sensibilidad somática.
En las áreas corticales secundarias, denominadas corteza de asociación, se interconectan varias porciones de la corteza del mismo hemisferio o del contralateral.
ÁREAS DE ASOCIACIÓN
A) El área parietooccipitaltemporal abarca:
1) El área parietal posterior, que contiene las coordenadas espaciales para todas las partes del lado contrario del cuerpo y el espacio extrapersonal contralateral.
2) El área de comprensión del lenguaje, denominada área de Wernicke, situada en la circunvolución temporal superior.
3) El área de procesamiento inicial del lenguaje visual (lectura) en la circunvolución angular del lóbulo parietal inferior.
4) Un área para la nominación de los objetos, ubicada en la porción anterior del lóbulo occipital.
B) El área de asociación prefrontal opera en estrecha relación con las áreas motoras del lóbulo frontal, planificando patrones complejos y secuencias de movimiento. Gran parte de las aferencias provienen de la corteza de asociación parietooccipitaltemporal y su salida principal alcanza el núcleo caudado, donde continúa el procesamiento.
C) La corteza de asociación límbica abarca el polo anterior del lóbulo temporal, la cara ventral del lóbulo frontal y parte de la corteza cingular. Participa de los procesos complejos de la conducta emocional y de la motivación, como el hipotálamo, los cuerpos amigdalitis y el hipocampo.
D) El área de reconocimiento facial se encuentra en las caras ventromediales de los lóbulos occipital y temporal.
Las funciones interpretativas del área de Wernicke, la circunvolución angular y las áreas motoras frontales del habla se encuentran más desarrolladas en uno de los hemisferios, el dominante. El hemisferio izquierdo es el dominante en el 95% de las personas, al margen de que sean diestras o zurdas, no se entiende todavía como uno de los hemisferios adquiere dominancia.
Al área de Wernicke se le suele asignar una función interpretativa general, ya que su lesión impide la comprensión del lenguaje hablado o escrito, aún cuando una persona no presente ningún déficit auditivo y pueda leer las palabras escritas en una página.
De la misma manera, la lesión de la circunvolución angular (permaneciendo intacta el área de Wernicke) puede dejar indemne la capacidad de entender el lenguaje hablado, pero impide la comprensión de las palabras escritas, lo que se conoce como ceguera para las palabras.
FUNCIONES INTELECTUALES SUPERIORES DE LAS ÁREAS DE ASOCIACIÓN PREFRONTALES
La función de la corteza prefrontal es compleja y multifactorial, y se explica, normalmente, describiendo los déficits que sufren personas con lesiones importantes de esta corteza.
1.- Disminución de la agresividad y respuesta social inadecuada. Se reconoce mejor cuando las lesiones afectan a la cara ventral de la corteza prefrontal, el área de asociación límbica.
2.- Incapacidad de alcanzar los objetivos o ejecutar pensamientos secuenciales. La corteza prefrontal recoge información de varias dispersas del encéfalo para dar solución a los problemas, tanto si se trata de una respuesta motora como de otro tipo.
3.- La corteza prefrontal, como asiento de la memoria operativa. La capacidad de almacenar y clasificar las informaciones que deben aplicarse en la función resolutiva de problemas se describe como memoria operativa. Al reunir estas porciones de información almacenadas , se pueden elaborar pronósticos, planes de futuro, demorar respuestas mientras recabar más información, contemplar las consecuencias de los actos antes de realizarlos, correlacionar información de varias fuentes, y controlar los actos con arreglo a las leyes sociales o morales.
FUNCIÓN DEL CEREBRO EN LA COMUNICACIÓN: RECEPCIÓN Y EMISIÓN DEL LENGUAJE
Existen dos aspectos de la comunicación: la recepción del lenguaje (el lado sensitivo) y su emisión (la vertiente motora) . Algunas personas pueden oír o reconocer palabras escritas o habladas, pero no entienden su significado.
Ello se debe a una lesión en el área de Wernicke; este estado se conoce como afasia receptiva o sensorial, o simplemente afasia de Wernicke.
Si la lesión se extiende más allá de los confines del área de Wernicke, aparece una incapacidad total para la comunicación a través del lenguaje, que se denomina afasia global.
Si una persona puede formular el lenguaje verbal en su mente, pero no vocalizar la respuesta, se habla de afasia motora, que denota una lesión del área de Broca en el lóbulo frontal; este estado también se puede denominar afasia de Broca.
Las lesiones que dañan las áreas correspondientes del lenguaje del hemisferio no dominante determinan una aprosodia sensitiva (incapacidad para entender la cualidad emocional del habla) o motora (incapacidad para impartir un contenido emocional al habla).
FUNCIÓN DEL CUERPO CALLOSO Y DE LA COMISURA ANTERIOR PARA TRANSMITIR LOS PENSAMIENTOS, LOS RECUERDOS, EL APRENDIZAJE Y OTROS TIPOS DE INFORMACIÓN ENTRE LOS DOS HEMISFERIOS CEREBRALES.
El cuerpo calloso proporciona interconexiones abundantes para casi todas las áreas de los hemisferios cerebrales, con excepción de la porción anterior de los lóbulos temporales, que se comunican a través de la comisura anterior. Algunas de las conexiones funcionales más importantes mediadas por estos dos fascículos fibrosos son estas:
- El cuerpo calloso facilita la comunicación del área de Wernicke del hemisferio izquierdo con la corteza motora del hemisferio derecho. Cuando falta esta conexión, no resulta posible el movimiento del lado izquierdo del cuerpo ante una orden.
- La información visual y somatosensitiva del lado izquierdo del cuerpo alcanza el hemisferio derecho. Sin el cuerpo calloso, esta información sensitiva no se extendería hasta el área de Wernicke del hemisferio izquierdo. En consecuencia, esta información no se procesa en el área de Wernicke, y se hace caso omiso del lado izquierdo y del campo visual izquierdo.
- Sin el cuerpo calloso, solo la mitad izquierda del cerebro podría entender las palabras escritas y habladas. El lado derecho solo entiende la palabra escrita y no el lenguaje verbal. Sin embargo, las respuestas emocionales pueden afectar a ambos lados del cerebro ( y del cuerpo ) si la comisura anterior está intacta.
PENSAMIENTOS, COCIENCIA Y MEMORIA
Los sustratos neurales de los tres procesos del pensamiento, la conciencia y la memoria no se conocen todavía bien. La teoría holística propone que el pensamiento obedece a una estimulación en forma de patrones de la corteza cerebral, tálamo y sistema límbico; cada una de estas regiones confiere un carácter o cualidad particular el proceso.
MEMORIA: FUNCIONES DE LA FACILITACIÓN Y LA INHIBICIÓN SINÁPTICAS
Las memorias aparecen por cambios en la transmisión sináptica entre las neuronas a raíz de la actividad neura previa. Estos cambios generan vías nuevas, vías facilitadas o vías inhibidas que aparecen en los circuitos neurales pertinentes. Estas vías nuevas o alteradas se denominan huellas de memoria.
Nuestra mente se me inundada de información sensitiva, y una de las funciones esenciales del cerebro consiste en despreciar la información irrelevante o extraña a través de un proceso llamado habituación.
Es evidente que algunas memorias duran solo un segundo y otras se extienden a los largo de horas, días, meses o años. Por eso, se han descrito tres categorías de memorias:
- Memorias a corto plazo, que duran únicamente segundos o minutos, salvo que se transformen en una memoria a largo plazo.
- Memoria a medio plazo, que dura entre días y semanas, pero acaba desapareciendo.
- Memoria a largo plazo, que, una vez almacenada, se puede recuperar años más tarde o incluso permanecer toda la vida.
MEMORIA A CORTO PLAZO:
La memoria a corto plazo se ilustra por el recuerdo de un número de teléfono nuevo a los pocos segundos o minutos si se piensa en él. Se investigan diversas teorías acerca del sustrato de este mecanismo:
- Este tipo de memoria se debe a una actividad neura continua en un circuito reverberante.
- Ocurre por la activación de sinapsis de terminaciones presinápticas que, de forma característica, dan lugar a una facilitación o inhibición prolongadas.
- Acumulación de calcio en las terminaciones axónicas puede, finalmente, incrementar la salida sinóptica desde ese terminal.
MEMORIA A MEDIO PLAZO:
Esta memoria puede deberse a cambios químicos o físicos pasajeros de la membrana presináptica o postsináptica, que persisten entre unos minutos y varias semanas. Algunas de estas observaciones empíricas sobre estos mecanismos provienen de estudios con el caracol Aplysia..
La estimulación de un terminal facilitador, simultánea a la activación de otra aferencia sensitiva, hace que se libere serotonina en los lugares sinópticos del terminal sensitivo.
La estimulación de los receptores de serotonina activa la adenilcitociclasa del terminal sensitivo principal, con lo que se forma monofosfato de adenosina cíclico (AMPc), que causa una liberación de proteína cintas y fosforera una proteína que bloquea los canales de potasio del terminal sensitivo.
El descenso en la conductancia de potasio prolonga los potenciales de acción que alcanzan el terminal sensitivo, con lo que aumenta la entrada de calcio en el terminal y se acelera la liberación del neurotransmisor desde el terminal sensitivo, facilitándose la transmisión por dicha sinapsis.
MEMORIA A LARGO PLAZO:
Se cree que la memoria a largo plazo es consecuencia de cambios estructurales en la sinapsis, que potencias o suprimen la conducción de las señales.
Estos cambios estructurales abarcan:
- Un aumento en el número de los lugares de liberación de las vesículas sinápticas.
- Un aumento en el número de las vesículas sinápticas.
- Un aumento en el número de los terminales sintéticos.
- Cambios en la forma o en el número de las espinas postsinápticas.
CONSOLIDACIÓN DE LA MEMORIA
Para que las memorias se conserven a largo plazo han de consolidarse, es decir, deben iniciarse los cambios químicos o estructurales que dan lugar a la memoria a largo plazo.
Por lo general, se precisan de 5 a 0 min para una consolidación robusta se requiere 1h o más. El proceso de consolidación se basa, según se cree, en la iteración.
La repetición de una misma información, una y otra vez, dentro de la mente potencia la transferencia a largo plazo de la memoria a corto plazo.
Con el tiempo, los rasgos esenciales de la experiencia sensitiva se van fijando cada vez más en los almacenes de la memoria. Además, durante la consolidación las memorias se codifican en diversas categorías de información.
IMPORTANCIA DE DETERMINADOS COMPONENTES ESPECÍFICOS DEL CEREBRO EN EL PROCESO DE LA MEMORIA
Las lesiones del hipocampo determinan una amnesia anterógrada con incapacidad para crear o almacenar memorias nuevas. Las memorias establecidas antes de la lesión no se afectan; al parecer, la causa reside en que el hipocampo (y el núcleo dorsomedial del tálamo) está conectado con los denominados centros de castigo y remuneración.
La pérdida de memoria a largo plazo ocurre en lesiones del tálamo y, a veces, cuando se daña el hipocampo. Se ha formulado la hipótesis de que el tálamo puede formar parte del mecanismo de búsqueda y lectura en los almacenes de la memoria.
Esto hace pensar que los mecanismos de la memoria para las funciones se reparten en más de una región encéfalica.