
Cinco tipos básicos de receptores sensitvos:
• Los mecanorreceptores detectan la deformación física de la membrana del receptor o del tejido que lo rodea de forma inmediata.
• Los termorreceptores detectan los cambios ( calor o frío) en la temperatura del receptor.
• Los nocirreceptores detectan la presencia de daño físico o químico en el receptor o en el tejido que lo rodea.
• Los fotorreceptores (electromagnéticos) detectan la luz (fotones) que incide en la retina.
• Los quimiorreceptores son los responsables del gusto y del olfato, detectan los valores de 02 y C02 en la sangre y también la osmolalidad de los líquidos tisulares.
El receptor, una vez activado, inicia los potenciales de acción en la fibra sensitiva asociada, que luego vehicula dichos impulsos hasta la médula espinal en forma de una «línea marcada>> a través de un nervio periférico. Estos impulsos o potenciales de acción se asemejan en todas las fibras sensitivas y pueden desplegar diferencias cualitativas en su amplitud o frecuencia; no obstante, un potencial de acción desencadenado por un estímulo doloroso no se percibe como singular y distinto del potencial de acción iniciado por cualquier otro receptor o modalidad sensitiva.

Lo que permite diferenciar un tipo de sensibilidad de otra es el lugar del sistema nervioso al que llega o en el que termina la fibra. Cada fibra o grupo de neuronas unido por fibras sensitivas relacionadas se denomina «línea marcada».
Cuando el receptor es activado por el estímulo adecuado, genera una corriente local llamada potencial de receptor. Da igual si el estímulo es mecánico, químico o físico (calor, frío, luz), la transducción determina un cambio en la permeabilidad iónica de la membrana del receptor y, en consecuencia, un cambio en la diferencia de potencial a través de esa membrana. La máxima amplitud del potencial del receptor, de unos 100 mV, se alcanza cuando la permeabilidad de la membrana al sodio alcanza el máximo.
A medida que aumenta la intensidad de un estímulo, la frecuencia de los potenciales de acción subsiguientes suele elevarse. La amplitud del potencial del receptor puede cambiar sustancialmente con una modificación relativamente pequeña de la intensidad, para luego aumentar de forma mínima con un estímulo de intensidad superior.
Esta adaptación tiene lugar a través de dos mecanismos. En primer término, el estímulo puede modificar las propiedades fisicoquímicas del receptor; así, cuando se deforma inicialmente un corpúsculo de Pacini (y aumenta la permeabilidad de membrana), el líquido redistribuye la presión aplicada por sus laminillas concéntricas. Esta redistribución se refleja en un descenso de la permeabilidad de membrana y el potencial de receptor disminuye o se adapta. En segundo término, la propia fibra sensitiva puede experimentar una acomodación. Este proceso, aunque no se conoce muy bien, podría depender de una «inactivación» gradual de los canales de sodio con el paso del tiempo.
Los receptores de adaptación lenta siguen transmitiendo las señales, con una frecuencia que apenas varía mientras se mantenga el estímulo. Por este motivo, se denominan «receptores tónicos» y pueden vehicular la fuerza del estímulo durante largos períodos. Los receptores de adaptación rápida se activan solo cuando cambia la intensidad del estímulo, por lo que se denominan «receptores de velocidad» o «detectores de movimiento».

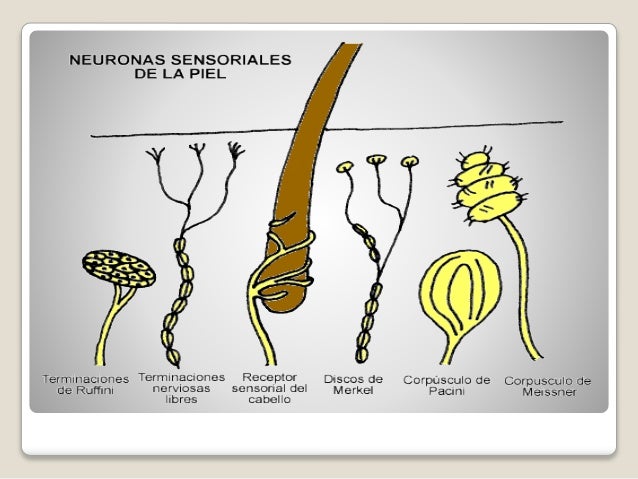
Normalmente, cada tronco sensitivo de un nervio periférico contiene varias fibras asociadas, cada una, a un número variable de receptores (más de 100 en el caso de las terminaciones nerviosas libres dela piel) en su terminación distal. El conjunto de todos los receptores y fibras de un mismo nervio define el campo receptor. Un estímulo intenso que se extienda por todo el campo receptor activaría todas las fibras del tronco sensitivo, y otro menos intenso, un número proporcionalmente menor. La graduación de la intensidad del estímulo depende de la participación de un número variable de fibras «paralelas» del mismo nervio (sumación espacial) o del cambio en la frecuencia con que los impulsos viajan por una misma fibra (sumación temporal).
Toda agrupación de neuronas, como la corteza cerebral, el tálamo o un núcleo concreto del tálamo, se puede denominar grupo neuronal. De manera característica, cada grupo neuronal posee un conjunto de varias aferencias (fibras de entrada), un campo receptor y una o varias «dianas» hacia las que se proyectan a través de una serie de axones eferentes organizados.
La estimulación liminar aumenta, como es evidente, el potencial de membrana por encima de los niveles de descarga de varias células, que generan potenciales de acción. En otras, el potencial de membrana se despolariza ligeramente, pero no lo suficiente para alcanzar el umbral (estimulación subliminal). Se dice que estas células se encuentran facilitadas, es decir, son más excitables, ya que se necesitan potenciales postsinápticos excitadores (PPSE) más pequeños de lo habitual para que la célula alcance el umbral y descargue potenciales de acción.
La divergencia de las señales de entrada es un rasgo común a algunos grupos neuronales. Esta divergencia puede adoptar una de dos formas. A través de un mecanismo de amplificación, la fibra de entrada puede ramificarse y entablar contacto con muchas neuronas del grupo; estas células postsinápticas se proyectan de forma unitaria sobre una diana o un número restringido de ellas. En el otro tipo de divergencia, las neuronas activadas del grupo se proyectan sobre dianas múltiples no relacionadas entre sí.
Un único axón eferente puede proporcionar una salida excitadora a una neurona del siguiente grupo (postsináptico) que por sí misma resulte excitadora (relevo), o bien establecer sinapsis con una interneurona inhibidora del siguiente grupo, que inhibiría, a su vez, las neuronas de relevo del grupo postsináptico. Esto se denomina circuito de inhibición recíproca.
El procesamiento de la señal en los grupos neuronales se basa a veces en un circuito reverberante u oscilatorio. Los axones de salida del grupo, en estos circuitos, emiten ramas colaterales que hacen sinapsis con interneuronas excitadoras situadas dentro del grupo. Estas interneuronas excitadoras retroalimentan las mismas neuronas de salida del grupo, determinando una secuencia autopropagadora de las señales. Los PPSE generados por las interneuronas excitadoras pueden facilitar o, en verdad, estimular la descarga por las neuronas de salida del grupo. Esta última situación constituye el sustrato de un grupo neuronal que emite un tren continuado de señales eferentes.
La conectividad extensa y diversa del sistema nervioso puede generar una inestabilidad funcional del cerebro cuando fracasan las operaciones. La crisis epiléptica es uno de los ejemplos más claros de esta inestabilidad. El sistema nervioso emplea dos mecanismos para combatir la inestabilidad funcional:
- El más destacado es la inhibición por retroalimentación
- El segundo método para limitar la inestabilidad se denomina fatíga sínáptica
