Esta es la primera entrada de mi nuevo blog. Acabo de empezar a escribir este nuevo blog. ¡Mantente al día para leer más entradas! Suscríbete a continuación para recibir notificaciones cuando publique nuevo contenido.
La cicatrización es el proceso de sanar los tejidos lesionados por medio de una compleja cascada de eventos celulares, los cuales son coordinados por mediadores solubles y conducen a la restitución física y funcional del tejido dañado. La cicatrización va a tener tres fases principales: hemostasia e inflamación, proliferación y remodelación. La literatura marca que, para fines didácticos, el proceso lo han dividido en esas tres fases, pero en la realidad se sobrelapan y evolucionan al mismo tiempo.
HEMOSTASIA Y FASE INFLAMATORIA
Considerada como la preparación de un sustrato o base orgánica y tisular que tiene como fin la curación y presupone una defensa contra otras lesiones o invasiones futuras, así como también la liberación de factores solubles quimiotácticos que controlan la permeabilidad de los vasos y otros que atraen o atrapan células
Plaquetas: Contienen al menos tres tipos de organelos: gránulos, cuerpos densos y lisosomas. Los gránulos contienen factores de crecimiento, así como factor transformador del crecimiento y fibronectina. Los cuerpos densos almacenan aminas vasoactivas (serotonina) y los lisosomas contienen proteasas. Las plaquetas liberan fibrinógeno, fibronectinas, trombospondina y factor de Von Willebrand. Interviniendo en la adhesión de las plaquetas a la colágena que quedó expuesta en la herida.
Coagulacion: El coágulo de fibrina no sólo tiene la función de hacer hemostasia, sino que junto con la fibronectina forma el armazón sobre el que migrarán los monocitos, fibroblastos y queratinocitos
Leucocitos: Los neutrófilos son las primeras células nucleadas en llegar. Esta migración es resultado de un proceso complejo que facilitan el paso de los neutrófilos a través de las células endoteliales de los capilares por medio de un mecanismo conocido como diapédesis. Para convertirse en cel de proceso inflamatorio.
FASE PROLIFERATIVA
La inflamación representa una función de limpieza y preparación, en tanto que la proliferación reconstruye, por lo que las fases no tienen una división cronológica y éstas ocurren de una manera conjunta y armónica, aunque, como en un incendio, no es posible reconstruir cuando todavía no se apaga el fuego. Al igual que la inflamación, la proliferación celular tiene elementos fundamentales.
La respuesta de las células epidérmicas inicia dentro de las primeras 24 horas de sufrida la lesión. A las 12 horas de perder contacto con sus homólogos vecinos. La mitosis tarda más en iniciar que la migración, por lo general de 48 a 72 horas después de la lesión. Las células se multiplican y movilizan hasta que entran en contacto con otras células epiteliales, momento en el cual se inhiben.
Angiogénesis: Los monocitos y los macrófagos producen factores que inducen la formación de nuevos vasos por los que se transporta oxígeno y nutrientes a la herida, y secretan sustancias biológicamente activa. Las células que intervienen en el proceso de angiogénesis son las endoteliales. Después se dividen, se forman vacuolas y se fusionan para crear un nuevo lumen
Fibroplasia y síntesis de colágeno: Los fibroblastos aparecen entre 48 y 72 horas después de ocasionada la herida, son estimulados y regulados por factores quimiotácticos complejos que provienen de los macrófagos presentes en la herida. Protagonistas de la proliferación y, junto con los nuevos vasos y la matriz, forman el tejido llamado de “granulación”
Contraccion de la herida: Disminución gradual del área de la herida por retracción de la masa central del tejido de granulación. Las fuerzas contráctiles producidas por este tejido son resultado de la acción de los miofibroblastos que contienen proteínas contráctiles.
FASE DE REMODELACIÓN
Cuando ha sido reparada la rotura de la continuidad de los tejidos, el estímulo angiógeno disminuye en intensidad y, al parecer, como respuesta a las tensiones elevadas de oxígeno en los tejidos se inicia un periodo en el que la herida madura, la cual presenta remodelación morfológica, también disminuyen la hiperemia y su vascularidad, asimismo se reorganiza el tejido fibroso neoformado.
CÉLULAS MADRE Y REGENERACIÓN DE LOS TEJIDOS
desde hace tiempo que en los organismos adultos y en el cordón umbilical de los recién nacidos existen células indiferenciadas, hoy se conoce que estas células conservan la habilidad de renovarse a sí mismas por mitosis celular y que son capaces de proliferar en la vida posnatal para producir o ser precursoras de estirpes celulares que se llegan a diferenciar y transformarse en células especializadas en respuesta a estímulos moleculares.
Las células mesenquimales que permanecen en estado indiferenciado reciben los nombres de células madre, células tallo, células troncales o células progenitoras (en inglés stem cells) y están identificadas como los actores de la renovación constante de los tejidos.
Transportar hormonas de una parte del cuerpo a otra.
Mantener un entorno apropiado en todos los líquidos tisulares del organismo.
El corazón actúa a modo de bomba, de modo que, al contraerse, genera la presión que impulsa a la sangre a través de una serie de vasos sanguíneos.
El ventrículo derecho bombea sangre hacia los pulmones, donde se oxigena la sangre; el ventrículo izquierdo bombea sangre oxigenada hacia todo el cuerpo.
La vía de sangre desde el corazón (ventrículo derecho), a través de los pulmones, y de regreso al corazón (aurícula izquierda) completa un circuito: la circulación pulmonar.
Las ramas arteriales provenientes de la aorta suministran sangre rica en oxígeno a todos los sistemas y, así, forman parte de la circulación sistémica.
COMPONENTES FUNCIONALES DE LA CIRCULACIÓN
El sistema circulatorio de los seres humanos consta de tres elementos funcionales básicos: una bomba (el corazón) que hace circular un líquido (la sangre) a través de una serie de contenedores (los vasos). Este sistema integrado es capaz de adaptarse a las circunstancias cambiantes de la vida normal.
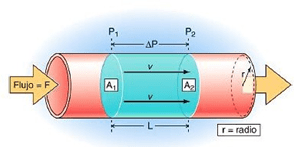
INTERRELACIONES E NTRE LA PRESIÓN, EL FLUJO Y LA RESISTENCIA
Por simplificar conceptos, pensemos en primer lugar en el corazón izquierdo como un generador de presión constante que mantiene una presión arterial media estable en su lugar de salida (es decir, en la aorta).
El flujo sanguíneo que atraviesa un vaso sanguíneo está determinado por dos factores:
1) Diferencia de presión de la sangre entre los dos extremos de un vaso, también denominado «gradiente de presión»
Los impedimentos que el flujo sanguíneo encuentra en el vaso, que se conoce como resistencia vascular.
La presión sanguínea se mide siempre como la diferencia de presión entre dos puntos
Los físicos miden la presión en unidades de gramo por centímetro cuadrado o dinas por centímetro cuadrado. Sin embargo, los fisiólogos suelen calibrar la presión arterial más a menudo por la altura a la que puede impulsar una columna de líquido.
Para comprender el flujo estable de la sangre impulsada por una presión de carga constante podemos aplicar las leyes de la hidrodinámica clásicas.
La ley más importante es la análoga a la ley de Ohm de la electricidad:
Es decir, la diferencia de presión (ΔP) entre un punto proximal, (presión P1) y uno distal (presión P2) es igual al producto del flujo, (F) por la resistencia (R) entre los dos puntos.
FLUJO DE SANGRE LAMINAR EN LOS VASOS
El flujo sanguíneo en el sistema cardiovascular es laminar o hidrodinámico. En el flujo laminar hay un perfil parabólico de la velocidad en el interior de un vaso sanguíneo, siendo la velocidad del flujo sanguíneo máxima en el centro del vaso y mínima hacia las paredes del vaso (El perfil parabólico se desarrolla porque la capa de sangre próxima a la pared del vaso se adhiere a esta última y, en esencia, no se mueve. La capa siguiente de sangre (hacia el centro) se desliza sobre la capa inmóvil y se desplaza un poco más rápido. Cada capa sucesiva de sangre hacia el centro se desplaza con mayor celeridad y con menor adherencia a las capas adyacentes.
FLUJO DE SANGRE TURBULENTO
Cuando aparece una irregularidad en un vaso sanguíneo. se desorganiza la corriente laminar y el flujo sanguíneo puede transformarse en turbulento. En un flujo turbulento, las corrientes del líquido no mantienen el perfil parabólico: en cambio, se mezclan radial y axialmente.
Dado que se consume energía para impulsar la sangre en sentido radial y axial, se necesita más energía
PRESIÓN SANGUÍNEA
Las presiones arteriales no son iguales a lo
largo de todo el sistema cardiovascular, si fuesen
iguales, la sangre no fluiría, ya que el flujo necesita que exista una fuerza de impulso (es decir, una diferencia de presión).
RESISTENCIA AL FLUJO SANGUÍNEO
La resistencia es el impedimento al flujo sanguíneo en un vaso, pero no se puede medir por medios directos. Por el contrario, la resistencia debe calcularse a partir de las determinaciones del flujo sanguíneo y de la diferencia de presión entre dos puntos del vaso.
LEY DE POISEUILLE
El flujo en un vaso ideal aumenta con la cuarta potencia del radio (ecuación de Poiseuille).
Hematocrito: proporción de sangre compuesta por eritrocitos. Si una persona tiene un hematocrito de 40 significa que el 40% del volumen sanguíneo está formado por las células y el resto es plasma. El hematocrito de un hombre adulto alcanza un promedio de 42, mientras que en las muy eres es de 38.
Estos valores son muy variables, dependiendo de si la persona tiene anemia, del grado de actividad corporal y de la altitud en la que reside la persona.
Después de que los alvéolos se hayan ventilado con aire limpio, la siguiente fase de la respiración es la difusión del oxígeno (O2) desde los alvéolos hacia la sangre pulmonar y la difusión del dióxido de carbono (CO2) en la dirección opuesta, desde la sangre a los alvéolos. El proceso de difusión es simplemente el movimiento aleatorio de moléculas en todas las direcciones a través de la membrana respiratoria y los líquidos adyacentes.
FÍSICA DE LA DIFUSIÓN GASEOSA Y PRESIONES PARCIALES DE GASES. BASE MOLECULAR DE LA DIFUSIÓN GASEOSA
Para que se produzca la difusión debe haber una fuente de energía. Esta fuente procede del movimiento cinético de las propias partículas. Excepto a la temperatura del cero absoluto, todas las moléculas de toda la materia están experimentando movimiento de manera continua. En el caso de las moléculas libres que no están unidas físicamente a otras, esto significa un movimiento lineal a una velocidad elevada hasta que chocan contra otras moléculas. Después rebotan en direcciones nuevas y siguen en movimiento hasta que chocan de nuevo con otras moléculas. De esta forma, las moléculas se mueven de manera rápida y aleatoria entre sí.
Difusión neta de un gas en una dirección: efecto de un gradiente de concentración
Si una cámara de gas o una solución tiene una concentración elevada de un gas particular en un extremo de la cámara y una concentración baja en el otro extremo, se producirá difusión neta del gas desde la zona de concentración elevada hacia la zona de concentración baja. La razón es evidente: hay muchas más moléculas en el extremo A de la cámara para difundir hacia el extremo B que moléculas para difundir en la dirección opuesta. Por tanto, las velocidades de difusión en cada una de las dos direcciones son diferentes proporcionalmente.
PRESIONES PARCIALES» DE GASES INDIVIDUALES
La presión está producida por múltiples impactos de partículas en movimiento contra una superficie. Por tanto, la presión de un gas que actúa sobre las superficies de las vías aéreas y de los alvéolos es proporcional a la suma de las fuerzas de los impactos de todas las moléculas de ese gas que chocan contra la superficie en cualquier momento dado. Esto significa que la presión es directamente proporcional a la concentración de las moléculas del gas.
Factores que determinan la presión parcial de un gas disuelto en un líquido
La presión parcial de un gas en una solución está determinada no solo por su concentración, sino también por el coeficiente de solubilidad del gas. Es decir, algunos tipos de moléculas, especialmente el CO2, son atraídas física o químicamente por las moléculas de agua, mientras que otros tipos de moléculas son repelidas. Cuando las moléculas son atraídas se pueden disolver muchas más sin generar un exceso de presión parcial en el interior de la solución. Por el contrario, en el caso de moléculas que son repelidas se generará una presión parcial elevada con menos moléculas disueltas.Estas relaciones se expresan mediante la fórmula siguiente, que es la ley de Henry.
LA DIFERENCIA DE PRESIÓN PROVOCA DIFUSIÓN DE GASES A TRAVÉS DE LÍQUIDOS
La difusión neta del gas desde la zona de presión elevada hacia la zona de presión baja es igual al número de moléculas que rebotan en esta dirección anterógrada menos el número que rebota en la dirección contraria, que es proporcional a la diferencia de presiones parciales de gas entre las dos zonas, denominada simplemente diferencia de presión para producir la difusión.
HUMIDIFICACIÓN DEL AIRE EN LAS VÍAS AÉREAS
El aire atmosférico está compuesto casi totalmente por nitrógeno y oxígeno; normalmente apenas contiene CO2 y poco vapor de agua. Sin embargo, tan pronto como el aire atmosférico entra en las vías aéreas está expuesto a los líquidos que recubren las superficies respiratorias. Incluso antes de que el aire entre en los alvéolos, se humidifica casi totalmente. La presión parcial de vapor de agua a una temperatura corporal normal de 37 °C es de 47 mmHg, que es, por tanto, la presión parcial de vapor de agua del aire alveolar. Como la presión total en los alvéolos no puede aumentar por encima de la presión atmosférica (760 mmHg a nivel del mar), este vapor de agua simplemente diluye todos los demás gases que están en el aire inspirado. La humidificación del aire diluye la presión parcial de oxígeno al nivel del mar desde un promedio de 159 mmHg en el aire atmosférico a 149 mmHg en el aire humidificado, y diluye la presión parcial de nitrógeno desde 597 a 563 mmHg.
EL AIRE ALVEOLAR SE RENUEVA LENTAMENTE POR EL AIRE ATMOSFÉRICO.
En promedio la capacidad residual funcional de los pulmones (el volumen de aire que queda en los pulmones al final de una espiración normal) en un hombre mide aproximadamente 2.300 ml. Sin embargo, solo 350 ml de aire nuevo entran en los alvéolos en cada inspiración normal y se espira esta misma cantidad de aire alveolar. Por tanto, el volumen de aire alveolar que es sustituido por aire atmosférico nuevo en cada respiración es de solo 1/7 del total, de modo que son necesarias múltiples inspiraciones para intercambiar la mayor parte del aire alveolar.
Importancia de la sustitución lenta del aire alveolar. La sustitución lenta del aire alveolar tiene una importancia particular en la prevención de cambios súbitos de las concentraciones de gases en la sangre. Esto hace que el mecanismo de control respiratorio sea mucho más estable de lo que sería de otro modo, y ayuda a prevenir los aumentos y disminuciones excesivos de la oxigenación tisular, de la concentración tisular de CO2 y del pH tisular cuando se produce una interrupción temporal de la respiración.
CONCENTRACIÓN Y PRESIÓN PARCIAL DE OXÍGENO EN LOS ALVÉOLOS.
El oxígeno se absorbe continuamente desde los alvéolos hacia la sangre de los pulmones, y continuamente se respira O2 nuevo hacia los alvéolos desde la atmósfera. Cuanto más rápidamente se absorba el O2, menor será su concentración en los alvéolos; por el contrario, cuanto más rápidamente se inhale nuevo O2 hacia los alvéolos desde la atmósfera, mayor será su concentración. Por tanto, la concentración de O2 en los alvéolos, y también su presión parcial, está controlada por: 1) la velocidad de absorción de O2 hacia la sangre, y 2) la velocidad de entrada de O2 nuevo a los pulmones por el proceso ventilatorio.
EL AIRE ESPIRADO ES UNA COMBINACIÓN DE AIRE DEL ESPACIO MUERTO Y AIRE ALVEOLAR
La composición global del aire espirado está determinada por: la cantidad del aire espirado que es aire del espacio muerto y la cantidad que es aire alveolar. La primera porción de este aire, el aire del espacio muerto de las vías aéreas respiratorias, es aire humidificado típico. Después cada vez más aire alveolar se mezcla con el aire del espacio muerto hasta que finalmente se ha eliminado el aire del espacio muerto y solo se espira aire alveolar al final de la espiración. Por tanto, el método para obtener aire alveolar para su estudio es simplemente obtener una muestra de la última porción del aire espirado después de que una espiración forzada haya eliminado todo el aire del espacio muerto.
La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las adventicias de las arterias y venas pulmonares. Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión solo ligeramente inferior a la presión aórtica. La circulación de alto flujo y baja presión que suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2). La arteria pulmonar recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
La arteria pulmonar se extiende solo 5 cm más allá de la punta del ventrículo derecho y después se divide en las ramas principales derecha e izquierda, que vascularizan los dos pulmones correspondientes. La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico. Esta gran distensibilidad permite que las arterias pulmonares se acomoden al gasto del volumen sistólico del ventrículo derecho. Las venas pulmonares, al igual que las arterias pulmonares, también son cortas. Drenan inmediatamente la sangre que les llega hacia la aurícula izquierda. La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho. La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg, valores que son solo un quinto de los del ventrículo izquierdo. Durante la sístole la presión en la arteria pulmonar es esencialmente igual a la presión que hay en el ventrículo derecho. después del cierre de la válvula pulmonar al final de la sístole, la presión ventricular cae súbitamente, mientras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones. la presión arterial pulmonar sistólica se sitúa normalmente en promedio en unos 25 mmHg en el ser humano, la presión arterial pulmonar diastólica es de aproximadamente 8 mmHg y la presión arterial pulmonar media es de 15 mmHg. La presión capilar pulmonar media es de 7mmHg. La presión media en la aurícula izquierda y en las venas pulmonares principales es en promedio de aproximadamente 2 mmHg en el ser humano en decúbito, y varía desde un valor tan bajo como 1 mmHg hasta uno tan elevado como 5 mmHg.
Se puede estimar la presión auricular izquierda con una exactitud moderada midiendo la denominada presión de enclavamiento pulmonar. Esta medida se consigue introduciendo un catéter en primer lugar a través de una vena periférica hasta la aurícula derecha, después a través del lado derecho del corazón y a través de la arteria pulmonar hacia una de las pequeñas ramas de la arteria pulmonar, y finalmente empujando el catéter hasta que se enclava firmemente en la rama pequeña.
El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares. La insuficiencia del lado izquierdo del corazón o el aumento de la resistencia al flujo sanguíneo a través de la válvula mitral como consecuencia de una estenosis mitral o una insuficiencia mitral hace que la sangre quede estancada en la circulación pulmonar, aumentando a veces el volumen de sangre pulmonar hasta un 100% y produciendo grandes aumentos de las presiones vasculares pulmonares. Dado que el volumen de la circulación sistémica es aproximadamente nueve veces el de la circulación pulmonar, el desplazamiento de sangre desde un sistema hacia el otro afecta mucho al sistema pulmonar, pero habitualmente tiene solo efectos circulatorios sistémicos leves.
Los factores que controlan el gasto cardíaco también controlan el flujo sanguíneo pulmonar. Para que se produzca una aireación adecuada de la sangre, esta debe distribuirse a los segmentos de los pulmones en los que los alvéolos estén mejor oxigenados.
La dinámica del intercambio de líquido a través de las membranas capilares pulmonares es cualitativamente la misma que en los tejidos periféricos. Sin embargo, cuantitativamente hay diferencias importantes, como se señala a continuación: 1. La presión capilar pulmonar es baja, de aproximadamente 7 mmHg, en comparación con una presión capilar funcional mucho mayor en los tejidos periféricos, de aproximadamente 17 mmHg. 2. La presión del líquido intersticial del pulmón es ligeramente más negativa que en el tejido subcutáneo periférico. (Esta presión se ha medido de dos formas: con una micropipeta insertada en el intersticio pulmonar, que da un valor de aproximadamente –5 mmHg, y midiendo la presión de absorción de líquido desde los alvéolos, que da un valor de aproximadamente –8 mmHg.) 3. La presión coloidosmótica del líquido intersticial pulmonar es de aproximadamente 14 mmHg, en comparación con menos de la mitad de este valor en los tejidos periféricos. 4. Las paredes alveolares son muy delgadas, y el epitelio alveolar que recubre las superficies alveolares es tan débil que se puede romper si la presión positiva en los espacios intersticiales es mayor que la presión del aire alveolar (>0 mmHg), lo que permite el paso de líquido desde los espacios intersticiales hacia los alvéolos.
La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las adventicias de las arterias y venas pulmonares. Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión solo ligeramente inferior a la presión aórtica. La circulación de alto flujo y baja presión que suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2). La arteria pulmonar recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
La arteria pulmonar se extiende solo 5 cm más allá de la punta del ventrículo derecho y después se divide en las ramas principales derecha e izquierda, que vascularizan los dos pulmones correspondientes. La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico. Esta gran distensibilidad permite que las arterias pulmonares se acomoden al gasto del volumen sistólico del ventrículo derecho. Las venas pulmonares, al igual que las arterias pulmonares, también son cortas. Drenan inmediatamente la sangre que les llega hacia la aurícula izquierda. La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho. La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg, valores que son solo un quinto de los del ventrículo izquierdo. Durante la sístole la presión en la arteria pulmonar es esencialmente igual a la presión que hay en el ventrículo derecho. después del cierre de la válvula pulmonar al final de la sístole, la presión ventricular cae súbitamente, mientras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones. la presión arterial pulmonar sistólica se sitúa normalmente en promedio en unos 25 mmHg en el ser humano, la presión arterial pulmonar diastólica es de aproximadamente 8 mmHg y la presión arterial pulmonar media es de 15 mmHg. La presión capilar pulmonar media es de 7mmHg. La presión media en la aurícula izquierda y en las venas pulmonares principales es en promedio de aproximadamente 2 mmHg en el ser humano en decúbito, y varía desde un valor tan bajo como 1 mmHg hasta uno tan elevado como 5 mmHg.
Se puede estimar la presión auricular izquierda con una exactitud moderada midiendo la denominada presión de enclavamiento pulmonar. Esta medida se consigue introduciendo un catéter en primer lugar a través de una vena periférica hasta la aurícula derecha, después a través del lado derecho del corazón y a través de la arteria pulmonar hacia una de las pequeñas ramas de la arteria pulmonar, y finalmente empujando el catéter hasta que se enclava firmemente en la rama pequeña.
El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares. La insuficiencia del lado izquierdo del corazón o el aumento de la resistencia al flujo sanguíneo a través de la válvula mitral como consecuencia de una estenosis mitral o una insuficiencia mitral hace que la sangre quede estancada en la circulación pulmonar, aumentando a veces el volumen de sangre pulmonar hasta un 100% y produciendo grandes aumentos de las presiones vasculares pulmonares. Dado que el volumen de la circulación sistémica es aproximadamente nueve veces el de la circulación pulmonar, el desplazamiento de sangre desde un sistema hacia el otro afecta mucho al sistema pulmonar, pero habitualmente tiene solo efectos circulatorios sistémicos leves.
Los factores que controlan el gasto cardíaco también controlan el flujo sanguíneo pulmonar. Para que se produzca una aireación adecuada de la sangre, esta debe distribuirse a los segmentos de los pulmones en los que los alvéolos estén mejor oxigenados.
La dinámica del intercambio de líquido a través de las membranas capilares pulmonares es cualitativamente la misma que en los tejidos periféricos. Sin embargo, cuantitativamente hay diferencias importantes, como se señala a continuación: 1. La presión capilar pulmonar es baja, de aproximadamente 7 mmHg, en comparación con una presión capilar funcional mucho mayor en los tejidos periféricos, de aproximadamente 17 mmHg. 2. La presión del líquido intersticial del pulmón es ligeramente más negativa que en el tejido subcutáneo periférico. (Esta presión se ha medido de dos formas: con una micropipeta insertada en el intersticio pulmonar, que da un valor de aproximadamente –5 mmHg, y midiendo la presión de absorción de líquido desde los alvéolos, que da un valor de aproximadamente –8 mmHg.) 3. La presión coloidosmótica del líquido intersticial pulmonar es de aproximadamente 14 mmHg, en comparación con menos de la mitad de este valor en los tejidos periféricos. 4. Las paredes alveolares son muy delgadas, y el epitelio alveolar que recubre las superficies alveolares es tan débil que se puede romper si la presión positiva en los espacios intersticiales es mayor que la presión del aire alveolar (>0 mmHg), lo que permite el paso de líquido desde los espacios intersticiales hacia los alvéolos.
La respiración proporciona oxígeno a los tejidos y retira el dióxido de carbono.
Las cuatro Funciones principales de la respiración son: 1) ventilación pulmonar, que se refiere al flujo de entrada y salida de aire entre la atmosfera y los alveolos pulmonares; 2) difusión de oxígeno y de dióxido de carbono entre los alveolos y la sangre; 3) transporte de oxígeno y de dióxido de carbono en la sangre y los líquidos corporales hacia las células de los tejidos corporales y desde las mismas, y 4) regulación de la ventilación y otras facetas de la respiración. Este capítulo analiza la ventilación pulmonar y los cinco capítulos posteriores abordan las otras funciones respiratorias más la fisiología de alteraciones respiratorias especiales.
MECÁNICA DE LA VENTILACIÓN PULMONAR.
MUSCULOS QUE CAUSAN LA EXPANSIÓN Y CONTRACCION PULMONAR.
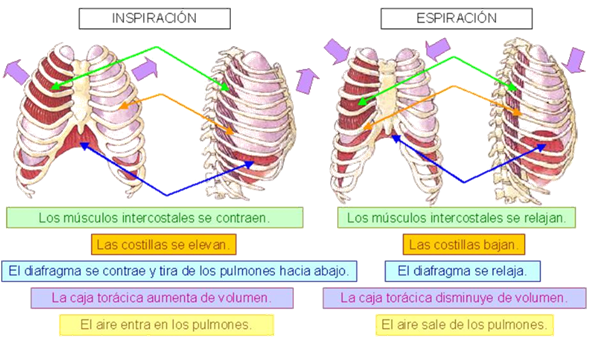
Los pulmones se pueden expandir y contraer de dos maneras: 1) mediante el movimiento hacia abajo y hacia arriba del diafragma para alargar o acortar la cavidad toracica, y 2) mediante la elevacion y el descenso de las costillas para aumentar y reducir el diametro anteroposterior de la cavidad toracica. muestra estos dos mecanismos. La respiracion tranquila normal se consigue casi totalmente por el primer mecanismo, es decir, por el movimiento del diafragma. Durante la inspiracion la contraccion del diafragma tira hacia abajo de las superficies inferiores de los pulmones. Despues, durante la espiracion el diafragma simplemente se relaja, y el retroceso elastico de los pulmones, de la pared toracica y de las estructuras abdominales comprime los pulmones y expulsa el aire. Sin embargo, durante la respiracion forzada las fuerzas elasticas no son suficientemente potentes para producir la espiracion rapida necesaria, de modo que se consigue una fuerza adicional principalmente mediante la contraccion de los musculos abdominales, que empujan el contenido abdominal hacia arriba contra la parte inferior del diafragma, comprimiendo de esta manera los pulmones.
El segundo metodo para expandir los pulmones es elevar la caja toracica. Esto expande los pulmones porque, en la posicion de reposo natural, las costillas estan inclinadas hacia abajo, como se muestra en el lado izquierdo de la figura 37-1, lo que permite que el esternon se desplace hacia abajo y hacia atras hacia la columna vertebral. Sin embargo, cuando la caja costal se eleva, las costillas se desplazan hacia adelante casi en linea recta, de modo que el esternon también se mueve hacia delante, alejandose de la columna vertebral y haciendo que el diametro anteroposterior del torax sea aproximadamente un 20% mayor durante la inspiracion máxima que durante la espiracion. Por tanto, todos los musculos que elevan la caja toracica se clasifican como musculos inspiratorios y los musculos que hacen descender la caja torácica se clasifican como musculos espiratorios. Los musculos mas importantes que elevan la caja toracica son los intercostales externos, aunque otros musculos que contribuyen son: 1) los musculos esternocleidomastoideos, que elevan el esternon; 2) los serratos anteriores, que elevan muchas de las costillas, y 3) los escalenos, que elevan las dos primeras costillas. Los musculos que tiran hacia abajo de la caja costal durante la espiracion son principalmente 1) los rectos del abdomen, que tienen el potente efecto de empujar hacia abajo las costillas inferiores al mismo tiempo que ellos y otros musculos abdominales tambien comprimen el contenido abdominal hacia arriba contra el diafragma, y 2) los intercostales internos. La figura 37-1 tambien muestra el mecanismo mediante el que actuan los intercostales externos e internos para producir la inspiracion y la espiracion. A la izquierda, durante la espiración las costillas estan anguladas hacia abajo, y los intercostales externos estan alargados hacia delante y hacia abajo. Cuando se contraen tiran de las costillas superiores hacia delante en relacion con las costillas inferiores y actuan como una palanca sobre las costillas para levantarlas hacia arriba, produciendo de esta manera la inspiracion. Los intercostales internos funcionan de manera exactamente opuesta, y actúan como musculos respiratorios porque se angulan entre las costillas en direccion opuesta y producen una palanca contraria.
PRESIONES QUE ORIGINAN EL MOVIMIENTO DE ENTRADA Y SALIDA DE AIRE DE LOS PULMONES.
El pulmon es una estructura elastica que se colapsa como un globo y expulsa el aire a traves de la traquea siempre que no haya ninguna fuerza que lo mantenga insuflado. Ademas, no
hay uniones entre el pulmon y las paredes de la caja torácica, excepto en el punto en el que esta suspendido del mediastino, la seccion media de la cavidad toracica, en el hilio. Por el contrario, el pulmon ≪flota≫ en la cavidad toracica, rodeado por una capa delgada de liquido pleural que lubrica el movimiento de los pulmones en el interior de la cavidad. Ademas, la aspiracion continua del exceso de liquido hacia los conductos linfaticos mantiene una ligera presion negativa entre la superficie visceral del pulmon y la superficie pleural parietal de la cavidad toracica. Por tanto, los pulmones estan sujetos a la pared toracica como si estuvieran pegados, excepto porque estan bien lubricados y se pueden deslizar libremente cuando el torax se expande y se contrae.
PRESIÓN PLEURAL Y SUS CAMBIOS DURANTE LA RESPIRACIÓN.
La presión pleural es la presión del líquido que está en el delgado espacio que hay entre la pleura pulmonar y la pleura de la pared torácica. Como se ha señalado antes, normalmente hay una aspiración ligera, lo que significa que hay una presión ligeramente negativa. La presión pleural normal al comienzo de la inspiración es de aproximadamente -5 cm H20, que es la magnitud de la aspiración necesaria para mantener los pulmones expandidos hasta su nivel de reposo. Después, durante la inspiración normal, la expansión de la caja torácica tira hacia fuera de los pulmones con más fuerza y genera una presión más negativa, hasta un promedio de aproximadamente -7,5 cm H20. Estas relaciones entre la presión pleural y las modificaciones del volumen pulmonar se muestran en la figura 37-2, en la que la parte inferior representa la negatividad creciente de la presión pleural desde – 5 hasta -7,5 durante la inspiración y la parte superior un aumento del volumen pulmonar de 0,5 l. Después, durante la espiración, se produce esencialmente una inversión de estos fenómenos.
PRESIÓN ALVEOLAR.
La presión alveolar es la presión del aire que hay en el interior de los alveolos pulmonares. Cuando la glotis está abierta y no hay flujo de aire hacia el interior ni el exterior de los pulmones, las presiones en todas las partes del árbol respiratorio, hasta los alveolos, son iguales a la presión atmosférica, que se considera que es la presión de referencia cero en las vías un movimiento de entrada de aire hacia los alveolos durante la inspiración, la presión en los alveolos debe disminuir hasta un valor ligeramente inferior a la presión atmosférica (debajo de cero). La segunda curva (denominada ≪presión alveolar≫) de la figura 37-2 muestra que durante la inspiración normal la presión alveolar disminuye hasta aproximadamente – l c m H20. Esta ligera presión negativa es suficiente para arrastrar 0,5 l de aire hacia los pulmones en los 2 s necesarios para una inspiración tranquila normal.
Durante la espiración se producen presiones contrarias: la presión alveolar aumenta hasta aproximadamente + lcm H20, lo que fuerza la salida del 0,5 l de aire inspirado desde los pulmones durante los 2 a 3 s de la espiración.
DISTENSIBILIDAD DE LOS PULMONES.
El volumen que se expanden los pulmones por cada aumento unitario de presión transpulmonar (si se da tiempo suficiente para alcanzar el equilibrio) se denomina distensibilidad p u lmonar. La distensibilidad pulmonar total de los dos pulmones en conjunto en el ser humano adulto normal es en promedio de aproximadamente 200 mi de aire por cada cm H20 de presion transpulmonar. Es decir, cada vez que la presión transpulmonar aumenta 1 cm H20 , el volumen pulmonar, despues de 10 a 20 s, se expande 200 ml.
Diagrama de distensibilidad de los pulmones. La figura 37-3 es un diagrama que relaciona los cambios del volumen pulmonar con los cambios de la presion transpulmonar. Observese que la relacion es diferente para la inspiración y para la espiracion. Cada una de las curvas se registra modificando la presion transpulmonar en escalones pequenos y permitiendo que el volumen pulmonar llegue a un nivel estable entre escalones sucesivos. Las dos curvas se denominan, respectivamente, la curva de distensibilidad inspiratoria y la curva de distensibilidad espiratoria y todo el diagrama se denomina diagrama de distensibilidad de los pulmones. Las caracteristicas del diagrama de distensibilidad están determinadas por las fuerzas elasticas de los pulmones. Estas se pueden dividir en dos partes: 1) fuerzas elasticas del tejido pulmonar en si mismo y 2) fuerzas elasticas producidas por la tension superficial del liquido que tapiza las paredes internas de los alveolos y de otros espacios aereos pulmonares. Las fuerzas elasticas del tejido pulmonar estan determinadas principalmente por las fibras de elastina y colágeno que estan entrelazadas entre si en el parenquima pulmonar. En los pulmones desinflados estas fibras estan en un estado
SURFACTANTE, TENSIÓN SUPERFICIAL Y COLAPSO DE LOS ALVÉOLOS.
Principio de la tension superficial. Cuando el agua forma una superficie con el aire, las moleculas de agua dela superficie del agua tienen una atraccion especialmente intensa entre si. En consecuencia, la superficie del agua siempre esta intentando contraerse. Esto es lo que mantiene unidas entre si las gotas de lluvia: una membrana muy contráctil de moleculas de agua que rodea toda la superficie de la gota de agua. Invirtamos ahora estos principios y veamos que ocurre en las superficies internas de los alveolos. Aquí la superficie de agua tambien intenta contraerse, lo que da lugar a un intento de expulsar el aire de los alveolos a través de los bronquios y, al hacerlo, hace que los alveolos intenten colapsarse. El efecto neto es producir una fuerza contráctil elastica de todos los pulmones, que se denomina fuerza elástica de la tension superficial.
El surfactante y su e fe c to sobre la tensión superficial. El surfactante es un agente activo de superficie en agua, lo que significa que reduce mucho la tension superficial del agua. Es secretado por celulas epiteliales especiales secretoras de surfactante denominadas celulas epiteliales alveolares de tipo II, que constituyen aproximadamente el 10% del área superficial de los alveolos. Estas celulas son granulares y contienen inclusiones de lipidos que se secretan en el surfactante hacia los alveolos. El surfactante es una mezcla compleja de varios fosfolipidos, proteinas e iones. Los componentes mas importantes son el fosfolipido dipalmitoilfosfatidilcolina, las apoproteinas del surfactante e iones calcio. La dipalmitoilfosfatidilcolina, junto a otros fosfolipidos menos importantes, es responsable de la reduccion de la tension superficial. Lo hace porque no se disuelve de manera uniforme en el liquido que tapiza la superficie alveolar, sino que parte de la molecula se disuelve, mientras
que el resto permanece sobre la superficie del agua en los alveolos. La tension de esta superficie es entre un doceavo y la mitad de la tension superficial de una superficie de agua pura. En terminos cuantitativos la tension superficial de diferentes liquidos en agua es aproximadamente la siguiente: agua pura, 72dinas/cm; los liquidos normales que tapizan los alveolos pero sin surfactante, 50dinas/cm; los liquidos normales que tapizan los alveolos con cantidades normales de surfactante incluidas, entre 5 y 30 dinas/cm.
EFECTO DE LA CAJA TORÁCICA SOBRE LA EXPANSIBILIDAD PULMONAR.
Hasta ahora hemos analizado la capacidad de expansion de los pulmones de manera aislada, sin considerar la caja toracica. La caja toracica tiene sus propias caracteristicas elásticas y viscosas, similares a las de los pulmones; incluso si los pulmones no estuvieran presentes en el torax, seguiría siendo necesario un esfuerzo muscular para expandir la caja toracica.
Distensibilidad del tórax y de los pulmones en conjunto.
La distensibilidad de todo el sistema pulmonar (los pulmones y la caja toracica en conjunto) se mide cuando se expanden los pulmones de una persona relajada o paralizada totalmente. Para hacerlo se introduce aire en los pulmones poco a poco mientras se registran las presiones y volumenes pulmonares. Para insuflar este sistema pulmonar total es necesario casi el doble de presion que para insuflar los mismos pulmones despues de extraerlos de la caja toracica. Por tanto, la distensibilidad del sistema pulmon-torax combinado es casi exactamente la mitad que la de los pulmones solos, 110 mi de volumen por cada cm H20 de presion para el sistema combinado, en comparacion con 200ml/cm H20 para los pulmones de manera aislada. Ademas, cuando los pulmones se expanden hasta alcanzar volumenes elevados o se comprimen hasta alcanzar volumenes bajos, las limitaciones del torax se hacen extremas; cuando se esta cerca de estos limites, la distensibilidad del sistema pulmon-torax combinado
puede ser menor de un quinto de la de los pulmones solos.
VOLUMENES Y CAPACIDADES PULMONARES
REGISTRO DE LAS VARIACIONES DEL VOLUMEN PULMONAR: ESPIROMETRIA
La ventilacion pulmonar puede estudiarse registrando el movimiento del volumen del aire que entra y sale de los pulmones, un metodo que se denomina espirometria. En la Sgura 37-5 se muestra un espirometro basico tipico. Esta formado por un tambor invertido sobre una camara de agua, con el tambor equilibrado por un peso. En el tambor hay ‘on gas respiratorio, habitualmente aire u oxigeno; un tubo conecta la boca con la camara de gas. Cuando se respira hacia el interior y el exterior de la camara, el tambor se eleva y desciende, y se hace un registro adecuado en una hoja de papel en movimiento.
ABREVIATURAS Y SIMBOLOS UTILIZADOS EN LAS PRUEBAS DE FUNCION RESPIRATORIA.
La espirometria es solo una de las muchas tecnicas de medición que utiliza a diario el neumologo. Muchas de estas técnicas de medida dependen mucho de calculos matematicos. Para simplificar estos calculos, asi como la presentacion de los datos de la funcion pulmonar, se han estandarizado diversas abreviaturas y simbolos. Los mas importantes se muestran en la tabla 37-1. Utilizando estos simbolos presentamos aqui algunos ejercicios algebraicos sencillos que muestran algunas de las interrelaciones entre los volumenes y capacidades pulmonares; el estudiante debe meditar y verificar estas interrelaciones.
VENTILACION ALVEOLAR
En ultimo termino, la funcion de la ventilacion pulmonar es renovar continuamente el aire de las zonas de intercambio gaseoso de los pulmones, en las que el aire esta proximo a la sangre pulmonar. Estas zonas incluyen los alveolos, los sacos alveolares, los conductos alveolares y los bronquiolos respiratorios. La velocidad a la que llega a estas zonas el aire nuevo se denomina ventilacion alveolar.
≪ESPACIO MUERTO≫ Y SU EFECTO SOBRE LA VENTILACION ALVEOLAR
Parte del aire que respira una persona nunca llega a las zonas de intercambio gaseoso, sino que simplemente llena las vías aereas en las que no se produce intercambio gaseoso, como la nariz, la faringe y la traquea. Este aire se denomina aire del espacio muerto, porque no es util para el intercambio gaseoso.
Durante la espiracion se expulsa primero el aire del espacio muerto, antes de que el aire procedente de los alveolos llegue a la atmosfera. Por tanto, el espacio muerto es muy desventajoso para retirar los gases espiratorios de los pulmones.
Medicion del volumen del espacio muerto. En el grafico de la figura 37-7 se presenta un metodo sencillo para medir el volumen del espacio muerto. Cuando se hace esta medicion, el paciente realiza subitamente una respiracion profunda de oxigeno. Esto llena todo el espacio muerto de oxigeno puro. Parte del oxigeno tambien se mezcla con el aire alveolar, aunque no sustituye completamente a este aire. Despues la persona espira a traves de un medidor de nitrogeno que registra rapidamente, y que hace el registro que se muestra en la figura. La primera porción del aire espirado procede de las regiones del espacio muerto de las vias aereas respiratorias, en las que el aire ha sido sustituido completamente por oxigeno. Por tanto, en la primera fase del registro solo aparece oxigeno, y la concentracion de nitrógeno es cero. Despues, cuando el aire alveolar comienza a llegar al medidor de nitrogeno, la concentracion de nitrogeno aumenta rapidamente porque el aire alveolar que contiene grandes cantidades de nitrogeno comienza a mezclarse con el aire del espacio muerto. Despues de que se haya espirado aun mas aire ya se ha eliminado todo el aire del espacio muerto de las vias aereas, ysolo queda aire alveolar. Por tanto, la concentración de nitrógeno que se registra alcanza una concentración de meseta igual a su concentración en los alveolos, como se muestra a la derecha de la figura. Pensándolo un poco el estudiante puede ver que la zona gris representa el aire que no tiene nitrógeno en su interior; esta area es una medida del volumen del aire del espacio muerto.
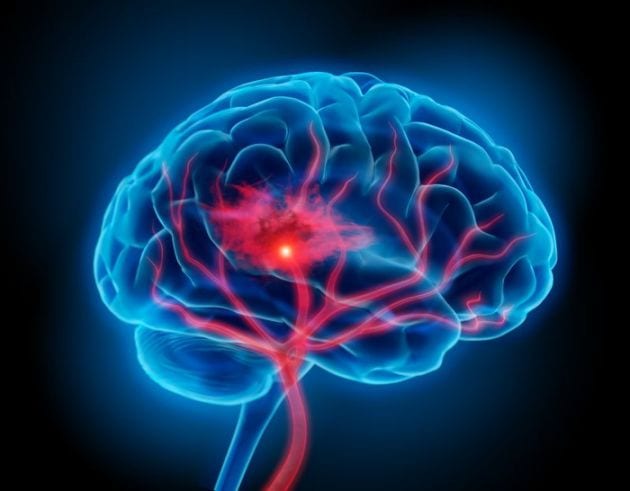
El flujo sanguíneo en el encéfalo es suministrado por cuatro grandes arterias, dos carotídeas y dos vertebrales, que se funden para formar el polígono de Willis en la base del encéfalo. Las arterias que parten del polígono de Willis se desplazan a lo largo de la superficie cerebral y dan origen a las arterias piales, que se ramifican en vasos más pequeños denominados arterias y arteriolas penetrantes.
Los vasos penetrantes están separados ligeramente del tejido encefálico por una extensión del espacio subaracnoideo denominada espacio de Virchow-Robin. Los vasos penetrantes se sumergen en el tejido encefálico, para dar lugar a arteriolas intracerebrales, que a su vez se ramifican en capilares en los que tiene lugar el intercambio entre la sangre y los tejidos de oxígeno, nutrientes, dióxido de carbono y metabolitos.
Regulación del flujo sanguíneo cerebral
Por término medio, el flujo sanguíneo normal a través del cerebro de una persona adulta es de 50 a 65 ml cada 100 g de tejido por minuto. Para todo el encéfalo, esta cantidad asciende 750 a 900 ml/min. Así pues, el encéfalo constituye únicamente en torno al 2% del peso corporal, pero recibe el 15% del gasto cardíaco en reposo. Al igual que sucede en la mayoría del resto de tejidos, el flujo sanguíneo cerebral está muy relacionado con el metabolismo tisular.
Según se cree, varios factores metabólicos contribuyen a la regulación del flujo sanguíneo cerebral:
1) la concentración de dióxido de carbono;
2) la concentración de iones hidrógeno,
3) la concentración de oxígeno, y
4) sustancias liberadas de los astrocitos, que son células no neuronales especializadas que parecen acompañar la actividad neuronal con la regulación del flujo sanguíneo local.
Importancia del control ejercido por el dióxido de carbono y los iones hidrógeno sobre el flujo sanguíneo cerebral
Una concentración alta de iones hidrógeno reduce mucho la actividad neuronal. Por tanto, es una suerte que su incremento también provoque un aumento del flujo sanguíneo, que a su vez retira del tejido cerebral iones hidrógeno, dióxido de carbono y otras sustancias formadoras de ácidos. La pérdida de dióxido de carbono elimina ácido carbónico de los tejidos; esta acción, junto con la extracción de otros ácidos, normaliza la concentración de iones hidrógeno.
Por tanto, dicho mecanismo sirve para mantener una concentración constante de iones hidrógeno en los líquidos cerebrales y ayuda así a conservar la actividad neuronal a un nivel normal y constante.
La falta de oxígeno como factor regulador del flujo sanguíneo cerebral
Los experimentos han demostrado que el descenso en la presión parcial de oxígeno (Po2) del tejido cerebral por debajo de unos 30 mmHg , comienza de inmediato a incrementar el flujo sanguíneo que recib; pues el funcionamiento cerebral sufre una perturbación a valores no mucho menores de la Po2, especialmente si llega a menos de 20 mmHg. A estos niveles tan bajos puede aparecer incluso un coma. Por tanto, el mecanismo de regulación local sobre el flujo sanguíneo cerebral por parte del oxígeno constituye una respuesta protectora muy importante contra el descenso de la actividad neuronal cerebral y, en consecuencia, contra cualquier trastorno en la capacidad mental.
Medición del flujo sanguíneo cerebral y sus efectos sobre la actividad cerebral
Se ha concebido un método para registrar el flujo sanguíneo en un mínimo de 256 segmentos aislados de la corteza cerebral humana al mismo tiempo. Para registrar el flujo sanguíneo en estos segmentos se inyecta en la arteria carótida una sustancia radiactiva, como xenón radiactivo; a continuación, se recoge la radiactividad de cada segmento cortical a medida que la sustancia atraviesa el tejido cerebral.
Con este fin, se ajustan 256 pequeños detectores de radiación contra la superficie de la corteza. La rapidez del ascenso y declive de la radiactividad en cada segmento tisular aporta una medida directa de la velocidad del flujo sanguíneo que lo atraviesa.
Microcirculación cerebral
Una característica estructural importante que presentan los capilares del encéfalo es que en su mayoría son menos «permeables» que los capilares sanguíneos casi de cualquier otro tejido del organismo. Una razón para este fenómeno radica en que cualquiera de sus caras se encuentra reforzada por los «podocitos neurogliales», que consisten en pequeñas prolongaciones procedentes de las células de la glía a su alrededor, que lindan con todas las superficies de los capilares y suministran un soporte físico para impedir su estiramiento excesivo en el caso de que suba demasiado la presión sanguínea capilar.
El «ictus» cerebral aparece cuando se obstruyen los vasos sanguíneos cerebrales. Casi todos los ancianos tienen bloqueadas algunas arterias pequeñas del encéfalo y hasta el 10% acaba sufriendo a la larga un bloqueo suficiente como para ocasionar un trastorno grave del funcionamiento cerebral, proceso llamado «ictus».
La mayoría de los ictus están causados por placas arterioescleróticas que aparecen en una o más de las arterias que irrigan el encéfalo.
Sistema del líquido cefalorraquídeo
Toda la cavidad que encierra el encéfalo y la médula espinal tiene una capacidad de unos 1.600 a 1.700 ml. De ellos, más o menos 150 ml están ocupados por el líquido cefalorraquídeo, y el resto por el encéfalo y la médula. Este líquido está presente en los ventrículos cerebrales, en las cisternas que rodean por fuera al encéfalo y en el espacio subaracnoideo alrededor del encéfalo y de la médula espinal. Todas estas cavidades se encuentran conectadas entre sí y la presión del líquido se mantiene a un nivel sorprendentemente constante.
Función amortiguadora del líquido cefalorraquídeo
Una función fundamental del líquido cefalorraquídeo consiste en amortiguar el encéfalo dentro de su bóveda sólida. El encéfalo y el líquido cefalorraquídeo poseen aproximadamente la misma densidad específica (tan solo difieren en un 4% más o menos), de modo que el encéfalo se limita a flotar en el seno del líquido. Por tanto, un golpe en la cabeza, si no es demasiado fuerte, desplaza todo el encéfalo a la vez que el cráneo, lo que evita que cualquier porción suya sufra una torsión transitoria por su acción.
Formación, flujo y absorción del líquido cefalorraquídeo
El líquido cefalorraquídeo se forma a una velocidad de unos 500 ml diarios, lo que supone el triple o el cuádruple de su volumen total en todo el sistema. Alrededor de dos tercios o más de esta cantidad se debe a la secreción desde los plexos coroideos en los cuatro ventrículos, sobre todo en los dos ventrículos laterales. Un poco más se produce en la superficie ependimaria de todos los ventrículos y en la aracnoides. Un pequeño porcentaje procede del encéfalo a través de los espacios perivasculares que quedan alrededor de los vasos sanguíneos que atraviesan el encéfalo.
La parte segregada en los ventrículos laterales pasa primero hacia el tercer ventrículo; después, tras la incorporación de una mínima cantidad más en esta cavidad, desciende a lo largo del acueducto de Silvio hacia el cuarto ventrículo, donde aún se añade otra minúscula proporción de líquido. Finalmente, sale del cuarto ventrículo por tres pequeños orificios, los dos agujeros laterales de Luschka y el agujero central de Magendie, para penetrar en la cisterna magna, un espacio de líquido que queda detrás del bulbo raquídeo y debajo del cerebelo. La cisterna magna se continúa con el espacio subaracnoideo que rodea al encéfalo y la médula espinal en su integridad. Casi todo el líquido cefalorraquídeo asciende a continuación desde la cisterna magna a través de estos espacios subaracnoideos alrededor del cerebro. Desde aquí, penetra por las múltiples vellosidades aracnoideas que sobresalen hacia el gran seno venoso sagital y otros senos venosos cerebrales, y las atraviesa. Por tanto, todo el líquido sobrante se vierte hacia la sangre venosa a través de los poros de estas vellosidades.
Presión del líquido cefalorraquídeo
La presión normal del sistema del líquido cefalorraquídeo en una persona tumbada en posición horizontal mide como promedio 130 mm de agua (10 mmHg), aunque esta presión puede bajar hasta 65 mm de agua o subir hasta 195 mm de agua incluso en una persona normal sana.
Barreras hematocefalorraquídea y hematoencefálica
Que separan la sangre del líquido cefalorraquídeo y del líquido encefálico, respectivamente. Hay barreras en los plexos coroideos y en las membranas de los capilares tisulares prácticamente en cualquier región del parénquima cerebral excepto en algunas zonas del hipotálamo, la glándula pineal y el área postrema, donde las sustancias difunden sin tantos problemas hacia los espacios tisulares.
En general, las barreras hematocefalorraquídea y hematoencefálica son muy permeables al agua, el dióxido de carbono, el oxígeno y la mayoría de las sustancias liposolubles, como el alcohol y los anestésicos; parcialmente permeables a electrolitos, como el sodio, el cloruro y el potasio, y casi totalmente impermeables a las proteínas plasmáticas y a la mayoría de las moléculas orgánicas grandes no liposolubles.
Edema cerebral
Una de las complicaciones más graves de las alteraciones dinámicas en el líquido cerebral. La causa más habitual de edema cerebral es el gran aumento de la presión en los capilares o la lesión de su pared, que la deja permeable al líquido. Un origen muy frecuente de este proceso es un golpe grave en la cabeza, que dé lugar a una conmoción cerebral, en la que tanto los tejidos como los capilares del cerebro quedan traumatizados. Una vez que comienza el edema cerebral, suele poner en marcha dos círculos viciosos:
El edema comprime los vasos, lo que a su vez reduce el flujo sanguíneo y produce una isquemia cerebral.
El descenso en el flujo sanguíneo cerebral también disminuye el aporte de oxígeno. Esto eleva la permeabilidad de los capilares, lo que permite un paso aún mayor de líquido
Metabolismo cerebral
Al metabolismo cerebral le corresponde aproximadamente el 15% del metabolismo total del organismo, aunque su masa no supone más que el 2% de la masa corporal íntegra. La principal necesidad metabólica neuronal consiste en bombear iones a través de sus membranas, sobre todo para transportar sodio y calcio al exterior de la membrana neuronal y potasio a su interior.
El encéfalo no es capaz de efectuar un gran metabolismo anaerobio. Una de las razones para ello estriba en el elevado índice metabólico de las neuronas, por lo que la mayor parte de la actividad neuronal depende de la liberación de oxígeno cada segundo desde la sangre.
En condiciones normales, casi toda la energía utilizada por las células del encéfalo llega suministrada por la glucosa extraída de la sangre. Un rasgo especial que caracteriza la liberación de la glucosa hacia las neuronas es que el transporte a través de la membrana celular no depende de la insulina, aunque su presencia sea necesaria para este proceso en la mayoría de las demás células del organismo. Por tanto, en los pacientes que tengan una diabetes grave con una secreción prácticamente nula de insulina, la glucosa aún difunde sin problemas hacia las neuronas, lo que es una gran suerte para evitar la pérdida de las funciones mentales en personas con diabetes.
El sistema nervioso autónomo es la porción del sistema nervioso que controla la mayoría de las funciones viscerales del cuerpo. Este componente interviene en la regulación de la presión arterial, la motilidad digestiva, las secreciones gastrointestinales, el vaciamiento de la vejiga urinaria, la sudoración, la temperatura corporal y otras muchas actividades. Algunas de ellas se encuentran casi del todo bajo su dominio en algunos casos y solo parcialmente en otros.
Organización general del sistema nervioso autónomo
El sistema nervioso autónomo se activa sobre todo a partir de centros situados en la médula espinal, el tronco del encéfalo y el hipotálamo. Asimismo, ciertas porciones de la corteza cerebral, sobre todo de la corteza límbica, pueden transmitir señales hacia los centros inferiores e influir de este modo en el control autónomo. El sistema nervioso autónomo también suele operar por medio de reflejos viscerales. Es decir, las señales sensitivas subconscientes procedentes de órganos viscerales pueden llegar a los ganglios autónomos, el tronco del encéfalo o el hipotálamo, y a continuación devolver unas respuestas reflejas subconscientes directamente a los órganos viscerales para controlar su actividad. Las señales autónomas eferentes se transmiten hacia los diversos órganos del cuerpo a través de sus dos componentes principales, denominados sistema nervioso simpático y sistema nervioso parasimpático, cuyas características y funciones se describen en el siguiente apartado.
Neuronas simpáticas preganglionares y posganglionares
Los nervios simpáticos son diferentes de los nervios motores esqueléticos por el hecho siguiente: cada vía simpática que se dirige desde la médula hasta el tejido estimulado está compuesta por dos células, una neurona preganglionar y una neurona posganglionar, a diferencia de la única neurona existente en la vía motora esquelética. El soma celular de cada neurona preganglionar está situado en el asta intermediolateral de la médula espinal; sus fibras van por una raíz ventral de la médula hasta llegar al nervio raquídeo correspondiente.
Nada más salir el nervio raquídeo del conducto raquídeo, las fibras simpáticas preganglionares lo abandonan y se encaminan a través de un ramo comunicante blanco hacia uno de los ganglios de la cadena simpática. Las fibras pueden seguir uno de los tres trayectos siguientes: 1) hacer sinapsis con neuronas simpáticas posganglionares en el ganglio al que llegan; 2) ascender o descender por la cadena y realizar sinapsis en cualquiera de los otros ganglios que la forman, o 3) recorrer una distancia variable a lo largo de la cadena y después irradiar hacia fuera a través de uno de los nervios simpáticos, para acabar haciendo sinapsis en un ganglio simpático periférico.
Anatomía fisiológica del sistema nervioso parasimpático
En el sistema nervioso parasimpático, se observa que las fibras parasimpáticas salen del sistema nervioso central a través de los pares craneales III, VII, IX y X; otras fibras parasimpáticas distintas abandonan la parte más inferior de la médula espinal por medio del segundo y el tercer nervio raquídeo sacro y, en ocasiones, por los nervios sacros primero y cuarto. En torno al 75% de todas las fibras nerviosas parasimpáticas están en el nervio vago (par craneal X), llegando a todas las regiones torácicas y abdominales del tronco. Por tanto, cuando un fisiólogo habla del sistema nervioso parasimpático muchas veces piensa sobre todo en los dos nervios vagos. Estos nervios suministran fibras parasimpáticas al corazón, los pulmones, el esófago, el estómago, todo el intestino delgado, la mitad proximal del colon, el hígado, la vesícula biliar, el páncreas, los riñones y las porciones superiores de los uréteres.
Neuronas parasimpáticas preganglionares y posganglionares
El sistema parasimpático, lo mismo que el simpático, posee neuronas preganglionares y posganglionares. Sin embargo, excepto en el caso de unos pocos nervios parasimpáticos craneales, las fibras preganglionares recorren sin interrupción todo el trayecto hasta el órgano que vayan a controlar. Las neuronas posganglionares están situadas en la pared del órgano. Las fibras preganglionares hacen sinapsis con estas neuronas, y unas fibras posganglionares extremadamente cortas, con una extensión que va desde una fracción de milímetro hasta varios centímetros de longitud, las abandonan para inervar los tejidos del órgano. Esta localización de las neuronas posganglionares parasimpáticas en el órgano visceral se aleja bastante de la organización de los ganglios simpáticos, debido a que los somas celulares de las neuronas posganglionares simpáticas casi siempre están situados en los ganglios de la cadena simpática o en otros ganglios aislados diferentes por el abdomen, en vez de hallarse en el órgano excitado.
Características básicas del funcionamiento simpático y parasimpático
Fibras colinérgicas y adrenérgicas: secreción de acetilcolina o de noradrenalina
Las fibras nerviosas simpáticas y parasimpáticas segregan básicamente una de las dos sustancias transmisoras de la sinapsis, acetilcolina o noradrenalina. Las fibras que liberan acetilcolina se llaman colinérgicas. Las que emiten noradrenalina se llaman adrenérgicas. Todas las neuronas preganglionares son colinérgicas tanto en el sistema nervioso simpático como en el parasimpático. La acetilcolina o las sustancias semejantes, al aplicarlas a los ganglios, excitarán las neuronas posganglionares tanto simpáticas como parasimpáticas. Todas o casi todas las neuronas posganglionares del sistema parasimpático también son colinérgicas. En cambio, la mayoría de las neuronas posganglionares simpáticas son adrenérgicas. Sin embargo, las fibras nerviosas simpáticas posganglionares dirigidas a las glándulas sudoríparas y, tal vez, a un número muy escaso de vasos sanguíneos son colinérgicas.
Receptores de los órganos efectores
Antes de que la acetilcolina, la noradrenalina o la adrenalina segregadas en una terminación nerviosa autónoma puedan estimular un órgano efector, primero deben unirse a sus receptores específicos en las células correspondientes. El receptor está situado en el exterior de la membrana celular, ligado como un grupo prostético a una molécula proteica que atraviesa toda la membrana celular. La fijación de la sustancia transmisora al receptor provoca un cambio de configuración en la estructura de la molécula proteica. A su vez, por regla general, la molécula modificada excita o inhibe a la célula: 1) causando un cambio en la permeabilidad de la membrana celular frente a uno o más iones, o 2) activando o inactivando una enzima ligada al otro extremo de la proteína receptora donde sobresale hacia el interior de la célula.
Dos tipos principales de receptores para la acetilcolina: receptores muscarínicos y nicotínicos
La acetilcolina activa sobre todo dos tipos de receptores, que reciben la denominación de receptores muscarínicos y nicotínicos. La razón de estos nombres radica en que la muscarina, un producto tóxico de las setas, solo activa los receptores muscarínicos y no los nicotínicos, mientras que la nicotina solo activa los nicotínicos. La acetilcolina estimula ambos. Los receptores muscarínicos, que usan proteínas G como mecanismo de señalización, están presentes en todas las células efectoras estimuladas por las neuronas colinérgicas posganglionares del sistema nervioso parasimpático, así como del sistema simpático. Los receptores nicotínicos son canales iónicos activados por ligando que se observan en los ganglios autónomos, a nivel de las sinapsis entre las neuronas preganglionares y las posganglionares de los sistemas simpático y parasimpático.
Receptores adrenérgicos: receptores α y β
También hay dos clases de receptores adrenérgicos; se denominan receptores α y receptores β. Existen dos tipos principales de receptores α, α1 y α2, que se unen a diferentes proteínas G. Los receptores β se dividen en receptores β1, β2 y β3 porque determinados productos químicos no actúan más que sobre alguno de ellos. Los receptores β también utilizan proteínas G para la señalización. La noradrenalina y la adrenalina, ambas segregadas a la sangre por la médula suprarrenal, poseen unos efectos un poco diferentes sobre la excitación de los receptores α y β. La noradrenalina estimula sobre todo los receptores α, pero también los receptores β, aunque en menor grado. La adrenalina activa ambos tipos de receptores aproximadamente por igual. Por tanto, los efectos relativos de la noradrenalina y la adrenalina sobre los diversos órganos efectores están determinados por los tipos de receptores que posean. Si todos son receptores β, la adrenalina será más eficaz en su acción excitadora.
Valor de la médula suprarrenal para el funcionamiento del sistema nervioso simpático
La adrenalina y la noradrenalina casi siempre se liberan de la médula suprarrenal al mismo tiempo que se excitan los diversos órganos directamente por la activación simpática generalizada. Por tanto, en realidad estas estructuras resultan estimuladas por dos vías: la directa a través de los nervios simpáticos y la indirecta a través de las hormonas de la médula suprarrenal. Los dos medios de estimulación se potencian entre sí y, en la mayoría de los casos, uno puede sustituir al otro. Por ejemplo, la destrucción de las vías simpáticas directas que van hacia los distintos órganos corporales no anula su excitación simpática debido a la noradrenalina y la adrenalina que todavía se liberan hacia la circulación sanguínea y producen una estimulación indirecta. En este mismo sentido, la desaparición de las dos médulas suprarrenales suele ejercer pocos efectos sobre el funcionamiento del sistema nervioso simpático debido a que las vías directas aún pueden realizar casi todas las tareas necesarias. Por tanto, el mecanismo doble de la estimulación simpática aporta un factor de seguridad, la sustitución de un método por otro en caso de que falte uno de ellos.
«Tono» simpático y parasimpático
Normalmente, los sistemas simpático y parasimpático están constantemente activos, y sus tasas basales de funcionamiento se conocen, respectivamente, como tono simpático y tono parasimpático. El valor de este factor reside en permitir que un solo sistema nervioso aumente o disminuya la actividad de un órgano estimulado. Por ejemplo, el tono simpático normalmente mantiene casi todas las arteriolas sistémicas contraídas más o menos hasta la mitad de su diámetro máximo. Si el grado de estimulación simpática aumenta por encima de su valor normal, estos vasos pueden contraerse aún más; por el contrario, si desciende por debajo de ese nivel, las arteriolas pueden dilatarse. Si no fuera por el tono simpático continuo de fondo, el sistema simpático solo sería capaz de ocasionar una vasoconstricción, nunca una vasodilatación.
Respuesta de «alarma» o de «estrés» en el sistema nervioso simpático
Cuando una gran porción del sistema nervioso simpático descarga a la vez (es decir, se produce una descarga masiva), esto aumenta por múltiples vías la capacidad del organismo para realizar una actividad muscular vigorosa de muchas formas, como se resume en la lista siguiente: 1. Aumento de la presión arterial. 2. Aumento del flujo sanguíneo para activar los músculos a la vez que disminuye la cantidad destinada a órganos como el tubo digestivo y los riñones, que no son necesarios para la actividad motora rápida. 3. Aumento de las tasas de metabolismo celular por todo el cuerpo. 4. Aumento de la concentración sanguínea de glucosa. 5. Aumento de la glucólisis hepática y muscular. 6. Aumento de la fuerza muscular. 7. Aumento de la actividad mental. 8. Aumento de la velocidad de coagulación sanguínea.
Control bulbar, pontino y mesencefálico del sistema nervioso autónomo
Muchas regiones neuronales pertenecientes a la formación reticular del tronco del encéfalo y situadas a lo largo del trayecto del fascículo solitario en el bulbo raquídeo, la protuberancia y el mesencéfalo, así como en múltiples núcleos especiales, regulan diversas funciones autónomas como la presión arterial, la frecuencia cardíaca, las secreciones glandulares en el tubo digestivo, el peristaltismo gastrointestinal y el grado de contracción de la vejiga urinaria. El control de cada una de ellas se estudia en el lugar correspondiente de este texto. Seguidamente se comentarán algunos de los factores más importantes controlados en el tronco del encéfalo son la presión arterial, la frecuencia cardíaca y la frecuencia respiratoria. En efecto, el corte transversal del tronco del encéfalo por encima de un nivel pontino medio permite mantener el control basal de la presión arterial sin cambios; pero impide su modulación por los centros nerviosos superiores, como el hipotálamo. Por el contrario, la sección inmediatamente por debajo del bulbo provoca su descenso hasta unos valores por debajo de la mitad de lo normal.
Según la explicación precedente, resulta evidente que la inyección intravenosa de noradrenalina produce básicamente los mismos efectos por todo el cuerpo que la estimulación simpática. Por tanto, la noradrenalina recibe el nombre de fármaco simpaticomimético o adrenérgico. La adrenalina y la metoxamina también son fármacos simpaticomiméticos, y hay otros muchos más. Estos compuestos difieren entre sí por el grado con el que estimulan los diferentes órganos efectores simpáticos y por la duración de su acción. En cuanto a este último aspecto, solo se extiende de 1 a 2 min en el caso de la noradrenalina y la adrenalina, mientras que dura de 30 min a 2 h en otros productos simpaticomiméticos diferentes de uso habitual. Los fármacos más importantes que estimulan unos receptores adrenérgicos específicos son la fenilefrina (receptores α), la isoprenalina o el isoproterenol (receptores β) y el salbutamol (solo receptores β2).
Fármacos que actúan sobre órganos efectores adrenérgicos: simpaticomiméticos
Fármacos que estimulan o bloquean las neuronas posganglionares simpáticas y parasimpáticas
Las neuronas preganglionares de los sistemas nerviosos simpático y parasimpático segregan acetilcolina en sus terminaciones, y esta acetilcolina estimula a su vez las neuronas posganglionares. Además, la inyección de acetilcolina también puede estimular las neuronas posganglionares de ambos sistemas, lo que genera al mismo tiempo efectos simpáticos y parasimpáticos por todo el organismo. Otro fármaco capaz de estimular las neuronas posganglionares de la misma manera que la acetilcolina es la nicotina, porque las membranas de todas estas neuronas contienen el receptor de acetilcolina de tipo nicotínico.
Por tanto, los productos que provocan efectos autónomos al estimular las neuronas posganglionares se llaman fármacos nicotínicos. Otros compuestos, como la metacolina, poseen acciones nicotínicas y muscarínicas, mientras que la pilocarpina solamente ejerce acciones muscarínicas. La nicotina excita las neuronas posganglionares simpáticas y parasimpáticas al mismo tiempo, lo que propicia una potente vasoconstricción simpática en los órganos abdominales y en las extremidades pero, a la vez, unos efectos parasimpáticos como el aumento de la actividad digestiva.
Todos nosotros somos conscientes de los múltiples estados posibles que presenta la actividad cerebral, como el sueño, la vigilia, la excitación extrema, e incluso los diversos estados de ánimo de una persona, entre ellos la euforia, la depresión y el miedo. Cualquiera de estos estados obedece a distintas fuerzas activadoras o inhibidoras generadas normalmente en el encéfalo.
SUEÑO
El sueño se define como el estado de inconsciencia del que puede ser despertada una persona mediante estímulos sensitivos o de otro tipo. Hay que distinguirlo del coma, que es el estado de inconsciencia del que no puede despertarse a una persona. El sueño está integrado por múltiples fases, desde el más ligero hasta el más profundo. Los investigadores que se dedican a este tema también lo dividen en dos tipos totalmente diferentes cuyas cualidades son distintas.
Dos tipos de sueño: de ondas lentas y de movimientos oculares rápidos (REM)
Cualquier persona atraviesa fases de dos tipos de sueño que alternan entre sí 1). Estos tipos reciben el nombre de: 1) sueño de movimientos oculares rápidos (sueño REM, por su denominación en inglés rapid eye movement), porque los ojos experimentan unos movimientos rápidos aun cuando la persona todavía está dormida, y 2) sueño de ondas lentas o no REM (NREM), en el que las ondas cerebrales son potentes y de baja frecuencia.
Sueño REM (sueño paradójico, sueño desincronizado)
El sueño REM se da en episodios que ocupan en torno al 25% del tiempo total en los jóvenes; estos episodios normalmente se repiten más o menos cada 90 min. Es un tipo de sueño no tan reparador y a menudo se asocia a sueños de gran viveza. La mayor parte del sueño durante la noche consiste en la modalidad de ondas lentas (NREM), que corresponde al sueño profundo y reparador que la persona experimenta durante la primera hora dormido después de haber estado despierto muchas horas.
Sueño de ondas lentas
La mayoría de nosotros podemos comprender las características del sueño profundo de ondas lentas si recordamos la última vez que estuvimos despiertos más de 24 h seguidas y a continuación el sueño profundo en el que caímos durante la primera hora después de acostarnos. Este sueño resulta sumamente reparador y va asociado a un descenso del tono vascular periférico y de otras muchas funciones vegetativas del cuerpo. Por ejemplo, se produce una disminución del 10 al 30% en la presión arterial, la frecuencia respiratoria y el índice metabólico basal. Aunque al sueño de ondas lentas se le llama a menudo «sueño sin sueños», durante su transcurso hay sueños y, en ocasiones, hasta pesadillas. La diferencia entre los sueños presentes en el sueño de ondas lentas y los que suceden en el sueño REM estriba en que estos últimos van asociados a una mayor actividad muscular del cuerpo. Además, los del sueño de ondas lentas no suelen recordarse porque no tiene lugar la consolidación de los sueños en la memoria.
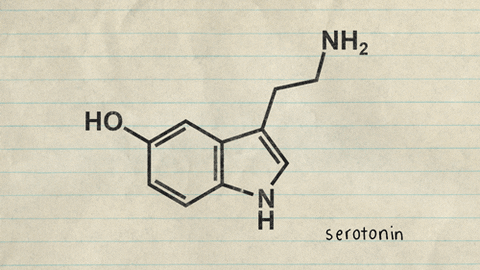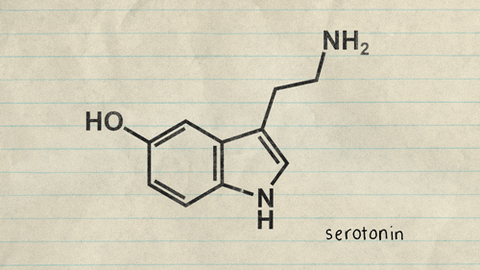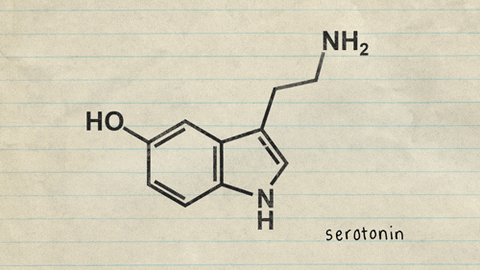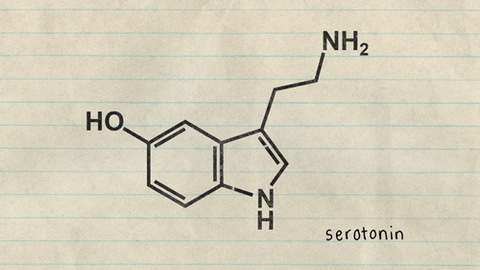
Centros nerviosos, sustancias neurohumorales y mecanismos capaces de causar sueño: posible función específica de la serotonina
a estimulación de diversas zonas específicas del encéfalo puede producir un sueño dotado de unas características próximas a las del sueño natural. Entre ellas figuran las siguientes: 1. La zona de estimulación para generar un sueño casi natural más constante son los núcleos del rafe en la mitad inferior de la protuberancia y en el bulbo raquídeo 2)La estimulación de algunas zonas en el núcleo del tracto solitario también puede generar sueño. Esta estructura es el punto de terminación en el bulbo raquídeo y en la protuberancia de las señales sensitivas viscerales que penetran a través de los nervios vago y glosofaríngeo. 3. El sueño puede promoverse mediante la estimulación de diversas regiones en el diencéfalo, como las siguientes: 1) la porción rostral del hipotálamo, sobre todo en el área supraquiasmática, y 2) en ciertas circunstancias una zona en los núcleos de proyección difusa del tálamo.
Ciclo de sueño y vigilia
Los comentarios precedentes meramente han identificado las zonas neuronales, los transmisores y los procesos relacionados con el sueño; no han explicado el funcionamiento cíclico recíproco que marca la sucesión entre la vigilia y el sueño; y por ahora tampoco existe ninguna explicación definitiva al respecto. Por tanto, podemos proponer el siguiente mecanismo posible como fuente del ciclo vigilia-sueño. Cuando los centros del sueño no están activos, los núcleos reticulares activadores del mesencéfalo y la parte superior de la protuberancia se encuentran liberados de su inhibición, lo que les permite una activación espontánea. Esta actividad espontánea a su vez excita a la corteza cerebral y al sistema nervioso periférico, los cuales devuelven numerosas señales de retroalimentación positiva a los mismos núcleos reticulares activadores para estimularles aún más. Por tanto, una vez que comienza la vigilia, su tendencia natural la lleva a mantenerse por sí sola debido a toda esta actividad de retroalimentación positiva.
El sueño tiene importantes funciones fisiológicas
El sueño produce dos tipos principales de acciones fisiológicas: en primer lugar, efectos sobre el sistema nervioso y, en segundo lugar, efectos sobre otros sistemas funcionales del cuerpo. Los efectos sobre el sistema nervioso parecen los más importantes con diferencia debido a que el cuerpo de cualquier persona cuya médula espinal haya sufrido una sección transversal a la altura del cuello (y, por tanto, carezca del ciclo vigilia-sueño por debajo del corte) no sufre unas consecuencias nocivas por debajo del nivel dañado que puedan atribuirse directamente al ciclo vigilia-sueño.
Ondas cerebrales
Los registros eléctricos recogidos en la superficie cerebral o incluso en la superficie de la cabeza ponen de manifiesto que existe una actividad eléctrica constante en el encéfalo. Tanto la intensidad como los patrones de esta variable vienen determinados por el grado de excitación que presentan sus diversos componentes como consecuencia del sueño, la vigilia o trastornos cerebrales como la epilepsia o incluso las psicosis.
Origen de las ondas α
Las ondas α no aparecerán en la corteza cerebral si no existen sus conexiones con el tálamo. En cambio, la estimulación de la capa inespecífica formada por el núcleo reticular que rodea al tálamo o de los núcleos «difusos» profundos en su interior a menudo produce ondas eléctricas en el sistema talamocortical a una frecuencia entre 8 y 13 por segundo, que corresponden a los valores naturales de las ondas α. Por tanto, se cree que las ondas α derivan de la oscilación de retroalimentación espontánea existente en este sistema talamocortical difuso, que tal vez abarca también el sistema reticular activador del tronco del encéfalo. Se supone que esta oscilación causa tanto la periodicidad de las ondas α como la activación sincrónica literalmente de millones de neuronas corticales durante cada onda.
Origen de las ondas δ
La sección transversal de los haces de fibras procedentes del tálamo hacia la corteza cerebral, que bloquea la activación talámica de esta estructura y elimina así las ondas α, no suprime en ella las ondas δ. Esto indica que puede haber cierto mecanismo de sincronización en el sistema neuronal cortical por sí solo, en esencia independiente de las estructuras inferiores en el cerebro, para dar origen a estas ondas δ.
Convulsiones y epilepsia
Las convulsiones son interrupciones temporales de la función encefálica causadas por una actividad neuronal excesiva e incontrolada. Según la distribución de las descargas neuronales, las manifestaciones de las convulsiones pueden estar comprendidas entre fenómenos experienciales apenas perceptibles y convulsiones espectaculares. Estas convulsiones sintomáticas temporales no suelen persistir si se corrige el trastorno subyacente. Pueden estar causadas por múltiples dolencias neurológicas o médicas, como trastornos agudos de electrólitos, hipoglucemia, fármacos (p. ej., cocaína), eclampsia, insuficiencia renal, encefalopatía hipertensiva, meningitis, y así sucesivamente. En torno al 5-10% de la población sufrirá al menos una convulsión en el curso de su vida.
A diferencia de las convulsiones sintomáticas, la epilepsia es una enfermedad crónica de convulsiones recurrentes que también puede oscilar entre síntomas breves y casi indetectables y períodos de vigorosa agitación y convulsiones. La epilepsia no es una enfermedad única. Sus síntomas clínicos son heterogéneos y reflejan múltiples causas subyacentes y mecanismos fisiopatológicos que provocan disfunción cerebral y lesiones, como traumatismos, tumores, infección o cambio.
Tratamiento de la epilepsia
La mayoría de los fármacos disponibles actualmente para tratar la epilepsia parecen bloquear el inicio o la extensión de las convulsiones, aunque no se conoce la forma exacta de acción de algunos de ellos, o bien tal vez realicen múltiples acciones. Algunos de los principales efectos de los diversos fármacos antiepilépticos son: 1) bloqueo de los canales del sodio dependientes del voltaje; 2) alteración de las corrientes de calcio ; 3) aumento en la actividad GABA ; 4) inhibición de los receptores de glutamato, el neurotransmisor excitador más común y 5) múltiples mecanismos de acción (p. ej., valproato y topiramato, que bloquean los canales del sodio dependientes del voltaje e incrementan los niveles de GABA en el encéfalo).
Esquizofrenia: posible funcionamiento excesivo de parte del sistema dopaminérgico
La esquizofrenia se manifiesta bajo numerosas variedades. Uno de los tipos más frecuentes se observa en la persona que oye voces y tiene delirios, un temor intenso u otras clases de sentimientos sin un origen real. Muchos esquizofrénicos sufren una gran paranoia, con una sensación de persecución a cargo de alguna fuente externa. Pueden presentar un lenguaje incoherente, disociación de ideas y secuencias anormales de pensamiento, y a menudo se encuentran retraídos, a veces adoptando una postura anormal e incluso rigidez.
Hay razones para pensar que la esquizofrenia tiene su origen al menos en una de las siguientes posibilidades: 1) múltiples áreas en los lóbulos prefrontales de la corteza cerebral cuyas señales nerviosas hayan quedado bloqueadas o en las que su procesamiento se vuelva disfuncional debido a que muchas sinapsis normalmente excitadas por el neurotransmisor glutamato pierdan su sensibilidad a esta sustancia; 2) una excitación excesiva de un grupo de neuronas que segreguen dopamina en los centros encefálicos del comportamiento, incluidos los lóbulos frontales, y/o 3) el funcionamiento anormal de un componente cerebral decisivo perteneciente al sistema límbico de control del comportamiento centrado en torno al hipocampo.
Enfermedad de Alzheimer: placas amiloides y pérdida de memoria
La enfermedad de Alzheimer se define como el envejecimiento prematuro del encéfalo, que suele comenzar al llegar a la mitad de la vida adulta y progresa con rapidez hasta una enorme pérdida de las capacidades mentales, semejante a la que se observa en las personas muy ancianas. Sus rasgos clínicos son los siguientes: 1) una afectación de la memoria de tipo amnésico; 2) un deterioro del lenguaje, y 3) un déficit visoespacial. Las alteraciones motoras y sensitivas, los trastornos de la marcha y las convulsiones son infrecuentes hasta las últimas fases de la enfermedad. Una observación constante en la enfermedad de Alzheimer es la desaparición neuronal en el componente de la vía límbica que se encarga del proceso de la memoria. La pérdida de esta función de la memoria tiene unas consecuencias devastadoras.
La enfermedad de Alzheimer es un trastorno neurodegenerativo de carácter progresivo y mortal que desemboca en una deficiencia de las capacidades de una persona para realizar sus actividades cotidianas, así como en una diversidad de síntomas neuropsiquiátricos y problemas del comportamiento durante las etapas finales de su evolución. Los pacientes con una enfermedad de Alzheimer suelen necesitar unos cuidados continuos en un plazo de pocos años desde su comienzo.
Las señales nerviosas del tronco del encéfalo activan el componente cerebral del encéfalo por dos caminos: 1) mediante la estimulación directa de un nivel de actividad neuronal y 2) por medio de la puesta en marcha de sistemas neurohormonales.
Control de la actividad cerebral mediante señales excitadoras continuas procedentes del tronco del encéfalo.
Área reticular excitadora del tronco del encéfalo.
El nivel de actividad del área excitadora en el tronco del encéfalo y, por tanto, el de todo el encéfalo, viene determinado en gran medida por la cantidad y el tipo de las señales sensitivas que llegan al encéfalo desde la periferia.
Aumento de la actividad del área excitadora ocasionado por las señales de retroalimentación que regresan desde la corteza cerebral. Este proceso sirve para mantener el nivel de activación cortical o incluso para potenciarlo.
El tálamo es un centro de distribución que controla la actividad en regiones específicas de la corteza
Un área reticular inhibidora se sitúa en la parte inferior del tronco del encéfalo. Se trata del área inhibidora reticular, que ocupa una posición medial y ventral en el bulbo raquídeo.
Control neurohormonal de la actividad encefálica.
Este mecanismo consiste en segregar sustancias hormonales neurotransmisoras excitadoras o inhibidoras sobre el parénquima del encéfalo. Contiene tres sistemas neurohormonales que se han estudiado con detalle en el encéfalo de la rata: 1) un sistema noradrenérgico; 2) un sistema dopaminérgico, y 3) un sistema serotoninérgico.
Sistemas neurohormonales en el encéfalo humano.
El locus ceruleus y el sistema de la noradrenalina.
La sustancia negra y el sistema de la dopamina.
Los núcleos del rafe y el sistema de la serotonina. Muchas de las neuronas que les componen segregan serotonina.
Las neuronas gigantocelulares del área excitadora reticular y el sistema de la acetilcolina.
Sistema límbico
La palabra «límbico» significa «limítrofe». En su origen, este término se empleó para describir las estructuras fronterizas que rodean a las regiones basales del cerebro; se refoere a todo el circuito neuronal que controla el comportamiento emocional y los impulsos de las motivaciones.
Anatomía funcional del sistema límbico; posición clave del hipotálamo.
Funciones de control vegetativo y endocrino del hipotálamo.
Esto regula las diferentes tipos de funciones.
1.Regulación cardiovascular 2.Regulación de la temperatura corporal 3.Regulación del agua corporal 4.Regulación de la contractilidad uterina y de la expulsión de leche por la mama 5.Regulación digestiva y de la alimentación 6.Control hipotalámico de la secreción de hormonas endocrinas por la adenohipófisis
Funciones conductuales a cargo del hipotálamo y de otras estructuras límbicas emparentadas con él.
Efectos ocasionados por la estimulación del hipotálamo
La estimulación del hipotálamo lateral no solo genera sed y ganas de comer,
La estimulación del núcleo ventromedial da lugar a unos efectos, sensación de saciedad, disminución del consumo de alimentos y tranquilidad.
La estimulación de una zona fina de los núcleos periventriculares, normalmente desemboca en temor y reacción frente al castigo.
El impulso sexual puede estimularse a partir de diversas zonas del hipotálamo, especialmente.
Efectos ocasionados por las lesiones hipotalámicas
Las lesiones en el hipotálamo lateral de ambos lados reducirán las ganas de beber y de comer casi a cero.
Las lesiones bilaterales de las regiones ventromediales del hipotálamo causan unos efectos ganas excesivas de beber y de comer, así como hiperactividad La estimulación o la lesión de otras regiones del sistema límbico.
Funciones de «recompensa» y de «castigo» cumplidas por el sistema límbico.
Estas cualidades afectivas también se denominan recompensa o castigo, o dicho de otro modo, satisfacción o aversión. La estimulación eléctrica de ciertas zonas límbicas agrada o satisface al animal, mientras que la actuación sobre otras regiones causa terror, dolor, miedo, reacciones de defensa o de huida, y todos los demás elementos acarreados por el castigo.
Importancia de la recompensa o el castigo en el aprendizaje y la memoria: habituación frente a refuerzo.
Cuando el estímulo sí causa una recompensa o un castigo en vez de la indiferencia, la respuesta de la corteza cerebral se vuelve progresivamente cada vez más intensa con su aplicación repetida en lugar de desvanecerse, y se dice que la respuesta está reforzada. Un animal robustece las huellas de memoria potentes para las sensaciones que sean gratificantes o penosas, pero, en cambio, adquiere una completa habituación frente a los estímulos sensitivos indiferentes.
Funciones específicas de otros componentes del sistema límbico
Funciones del hipocampo El hipocampo es la porción alargada de la corteza cerebral que se dobla hacia dentro para formar la cara ventral de gran parte del ventrículo lateral por su interior. Uno de sus extremos linda con los núcleos amigdalinos, y a lo largo de su borde lateral se fusiona con la circunvolución parahipocámpica, que es la corteza cerebral situada en la parte ventromedial de la cara externa del lóbulo temporal.
Función del hipocampo en el aprendizaje. Amnesia anterógrada después de la extirpación bilateral de los hipocampos Los pacientes que han sufrido la extirpación quirúrgica de porciones bilaterales de los hipocampos para el tratamiento de la epilepsia son capaces de recuperar satisfactoriamente la mayoría de los recuerdos aprendidos con antelación. No obstante, durante apenas un momento consiguen recordar lo que acontece en el curso de sus actividades. Este fenómeno, denominado amnesia anterógrada, se explicó en el.